Review Article - (2023) Volume 9, Issue 6
Received: 01-Jan-2025, Manuscript No. JFIM-22-68179;
Editor assigned: 04-Jan-2025, Pre QC No. JFIM-22-68179(PQ);
Reviewed: 18-Jan-2025, QC No. JFIM-22-68179;
Revised: 22-Jan-2025, Manuscript No. JFIM-22-68179(R);
Published:
30-Jan-2025
, DOI: 10.37421/2572-4134.2024.10.254
Citation: Tembo, Rostern. "The Mechanism of VDJ Recombination in the Human Immune System." J Food Ind Microbio 9 (2025): 254.
Copyright: �?�© 2025 Tembo R. This is an open-access article distributed under the terms of the creative commons attribution license which permits unrestricted
use, distribution and reproduction in any medium, provided the original author and source are credited.
During the maturation of every B cell and T cell in the body, there exists a process known as VDJ recombination, which acts to instate a drastic change in the cells’ genetic makeup. The purpose of the process is to completely randomize the way the genes are arranged so that the consequential immunoglobulins and T cell receptors can take on wildly different conformations, effectively preparing for the infinite possibilities of invasions that will inevitably occur. Taking only one of each of the numerous V, D, and J segments within a locus, proteins involved in the recombination function to bring said segments adjacent to one another, cleave off their ends, and attach them side by side via extra nucleotides placed in between them. As it is the process solely responsible for the body’s ability to protect and effectively memorize a plethora of different pathogens, antigens, and viruses, VDJ recombination almost single handedly defines the role of the human immune system at large. One feature of great importance in adaptive immunity is the ability to respond to an enormous number of different antigens.
The somatic assembly of the T Cell Receptor (TCR) genes generates a diverse T cell repertoire and is an essential component of the thymocytes development.
B cells • DNA • Immunoglobulin • T cell receptors • VDJ recombination • Recombination Signal Sequence (RSS)
The immune system is possibly one of the most intricate and important systems in the living organism. Normally unless one is ill, not a lot of attention is paid to the immune system. However, due to the recent global COVID-19 pandemic (2020-2022), the immune system has been given more attention than ever before. What makes up the immune system? The innate and the adaptive immune system (B and T cells) work independently as well as cohesively to combat foreign invaders such as bacteria, viruses, and fungi whose main aim is to cause harm and potentially death. To combat the said pathogens, the host body must have a repertoire of receptors found on the B and T cells whose sole purpose is to defend the body against pathogens. To do this, VDJ recombination of genes for the receptors’ diversity is a necessity. The VDJ recombination is a process of somatic recombination of gens that transpires solely in B and T cells during their maturation by randomly combing gene segments called Variable (V), Diversity (D), and Joining (J) genes. A highly diverse repertoire of antibodies and T cell receptors will be found in B and T cells, respectively, as a result [1]. VDJ recombination is fundamentally one of the most important processes our immune system carries out. Without VDJ recombination, there would be a weak adaptive immune system, and humanity would not stand a chance against the diverse repertoire of pathogens that constantly try to attack. Therefore, having a highly diverse repertoire of receptors is vital to our survival because these B Cells Receptors (BCRs) and T Cell Receptors (TCRs) can recognize virtually any foreign antigen [2].
The fundamental goal of every B cell and T cell is to acquire a specific, randomly selected genome that may be used to form their main immunological proteins. For B cells and their immediate successors, the plasma cells, are the immunoglobulins (or antibodies), whereas, for T cells, these are the T Cell Receptors (or TCRs). The latter is responsible for the recognition of foreign, potentially harmful particles antigens that have been bound to Major Histocompatibility molecules (MHC) on antigen presenting cells, whereas the former is used to bind to said particles to prohibit them from binding to their intended sites.
When this is the case, it is important for cells that protect the body against a multitude of highly diverse pathogens and infectious agents to have a highly diverse repertoire of both receptors and immunoglobulins. This fact highlights the need for a mechanism that can effectively recombine genes in the cells, which can then be transcribed and translated to form a wide variety of resulting molecules [3].
VDJ recombination archaically known as “antigen receptor gene rearrangement” fulfills this duty through a greatly randomized procedure. The VDJ recombination generates a highly diverse set of immunoglobulin and T cell receptor genes by the regulated joining of individual V,D and J gene segments. The reaction is initiated by two lymphocyte specific proteins called Recombination Activating Genes 1 and 2 (RAG1 and RAG2), that bind to the conserved Recombination Signal Sequences (RSSs) that frank all recombining gene segments in the immunoglobulin and T cell receptor loci [3]. As the B cells and T cells are facing the long, arduous task of maturation in the bone marrow and thymus, respectively, VDJ recombination, in a pathway believed to be evolutionarily inspired by transposition, succeeds in taking the genomes they received from their predecessors the lymphoid progenitors and restricting them by removing all but a few pieces of immunological information so that each cell can make a unique receptor or immunoglobulin [4,5]. These genomes, which are equipped with several each of V, J, and sometimes D sections, encode amino acid sequences that correspond to three adjacent sections of both the outer light chains and inner heavy chain on an immunoglobulin, as well as the outer variable regions on the chains for the T cell receptor [6]. Later, when an infection by a pathogen occurs, the randomized, one in a million cells that can efficiently recognize and/or bind to the pathogen are selected to proliferate and overwhelm the infection with their recombinant genomic products, beginning the action of the humoral immune system against the entirety of the infection. With all this in mind, the importance of the role that VDJ recombination plays in the human body cannot be understated.
Before B cells mature, the range of what their immunoglobulins can turn out to be after recombination can be found in three specific loci of their genomes; one dedicated to the heavy chain, located in humans on chromosome 14, and two dedicated to the light chains on either outer end of the Fab fragment, referred to as the λ (lambda) light chain, located on chromosome 2, and the κ (kappa) light chain, located on chromosome 22. For T cells, there are four loci that undergo recombination, those being for the α (alpha) chain and δ (delta) chain on chromosome 14, and the β (beta) chain and γ (gamma) chain on chromosome 7. Each of these loci contains a certain number of V (variable), D (diversity), and J (joining) segments hence the term “VDJ” recombination in addition to constant segments that do not undergo recombination and encode the parts of the immunoglobulin and T cell receptors that act as anchors to their respective cells [7]. It should be noted that only the heavy chain, β chain, and δ chain loci possess D segments in their sequences; the ones for the light chains, α chain, and γ chain have none, and, though the mechanism remains unchanged otherwise, they are said to only undergo “VJ” recombination instead. As such, VDJ recombination involves two recombination processes one that brings together the “D” and “J” segments and another that brings together the “V” with the newly formed “DJ” and VJ recombination only involves single recombination [8]. VDJ recombination is a process of somatic recombination that transpires solely in B and T cells during their maturation by randomly combining gene segments called Variable (V), Diversity (D), and Joining (J) genes. In simpler terms, V(D)J recombination chooses a pair of segments, then introduces a double strand, breaks adjacent to each segment, and then deletes or, in some cases, inverts the intervening DNA and then ligates the segments together.
Because it requires specific parts of the genome to be kept and others to be removed, the process utilizes a plethora of different enzymes, each of which is responsible for tweaking the DNA in its own way. As the procedure begins, the proteins are assembled to cut and splice the DNA such that the V, D, and J segments randomly selected to remain in the genome are brought together, cutting out most of what separated them. Each of the segments contains a “Recombination Signal Sequence,” (or RSS), on both ends that effectively act as cues that they are intended to be recombined by the enzymes, acting as docks for the enzymes to land in the correct position and orientation. All RSSs are organized with a heptamer (seven base pair) sequence on the side next to the segment, a nonamer (nine base pair) sequence on the other side, and a variable “spacer” in between them, which can either be twelve or twenty-three base pairs long. Depending on which locus they are located in, the two types of spacers are distributed to the V, D, and J segments differently; for example, in the B cell λ light chain locus, the 23 spacers are given to the V segments and the 12 spacers to the J segments, whereas the opposite is true for the κ light chain locus.
For enzymes to begin recombination, the specific segments randomly selected for V, D, and J must become adjacent to one another, forming a loop in the chromosome. This loop is held together by the first enzymes in the line of VDJ recombination: the “recombination activating gene,” or RAG, proteins, which come in pairs. The first, denoted RAG1, is responsible for recognizing the two gene sequences to dock on to and from the loop, while the second, RAG2, promotes stable binding and cleaving after docking. Other proteins that allow the DNA to bend far enough so that the process can occur, such as High Mobility Group Protein 1(HMG1), are required as well. Though the RSS spacers previously mentioned act to give enough room for these enzymes, they are also important for the 12/23 rule, which states that, in order for the segments to combine for recombination, a twelve base pair spacer can only become adjacent to a twenty-three-base pair spacer, and vice versa. For example, this means that for both the λ light chain and κ light chain loci, V segments can only align with J segments, whereas a V cannot align with another V. This rule also prohibits the possibility of a V segment from the λ light chain locus to accidentally cross-combine with a J segment from the κ light chain locus, which would form an undesired, dysfunctional amino-acid sequence [9].
In the DNA loop created by the RAG1-2 complex, there now exists a piece of DNA variable in length which holds most of the RSSs and may hold V, D, or J segments of its own which will go unused that will effectively be excised out of the genome by the RAG-2 protein via coupled cleavage. These are referred to as the “signal joints,” which, after their expulsion, are eventually degraded, whereas the rest of the genome left in two pieces, including the selected segments now adjacent to one another, are the “coding joints” [10]. The free ends of both the coding joints and the signal joints are unstable and are in need of stabilization by DNA repair enzymes named Ku70 and Ku80, starting the pathway of nonhomologous end joining. On the coding joints, these proteins will fuse the ends of the two separated nucleotide backbones into one via a transesterification reaction, forming covalent structures that are colloquially known as “hairpins;” on the signal joints, however, the ends are left blunt, as the two terminal sequences they receive from the RSSs are much easier to recombine due to the universal base pair organization of the heptamers and nonamers they contained. The next step is to bring the two coding joints together into one piece, breaking the hairpins and placing the segments together in sequence, which requires the help of DNA dependent protein kinase (DNA-PK) and the protein artemis, two other DNA repairing proteins that are combined to form the artemis complex.
These necessary incisions are made, leaving a short, palindromic sequence of unbound P nucleotides. Despite this, the ends of both sides of DNA are still likely unable to come together, as the pairs of backbones may be uneven or ragged from the cuts, which, if they annealed, would cause the resulting connected DNA to be uneven in base pairs. The next enzyme, named “Terminal Deoxynucleotidyl Transferase” (TDT), fixes this issue by adding random nucleotides, known as N nucleotides, to the backbones on both sides such that they will match. This protein’s actions can be reversed by an exonuclease if either the added nucleotides number too many or if certain base pairs are not compatible with each other, but no true order of action between the two processes is set in stone. Something important to note about this process is that the number of nucleotides added or subtracted from the original amount by either protein exponentially greatens the possibility of variability within what the eventual immunoglobulin or T cell receptor will become; this is because, in protein translation, one amino acid corresponds to three nucleotides on the mRNA strand, which are collectively called a codon. Therefore, if a net amount of extra nucleotides other than a multiple of three are put in the sequence by the TDT and exonuclease, the intended reading frame will not be preserved, leading to a completely different amino acid sequence as the end product. As an example, the DNA sequence TGG, which corresponds to the mRNA codon ACC and encodes for threonine, can be completely altered by just a single A being added before it (ATG), which would instead correspond to the codon UAC, encode alternatively for tyrosine, and cause the rest of the DNA sequence further along to be offset by one and completely alter the original set of translational instructions.
After this is achieved, one final step is required in the recombination the ligation of the strands. With the help of the protein XRCC4, DNA ligase IV is recruited to catalyze the end joining DNA double-strand break repair pathway, also removing any mismatching nucleotides. The ligase also ligates the separate ends of the signal joints of the excised piece of DNA without the need for any artemis or TDT proteins, which leaves behind a small ring of DNA permanently lost from the genome, destined for the signaling that the B cell or T cell has matured (where the joints get their name) and eventual degradation soon after. With ligation completed, VDJ recombination has been fully accomplished, and what results is an “imprecise” coding joint: The selected segments being side by side with only a small number of unrelated base pairs between them. Not long afterward, these newly formed DNA sequences will be transcribed and translated so that the cell’s specific immunoglobulins or T cell receptors can contribute to the body’s immune system; if their distinct conformations prove to be successful against an invasion, the cells will be nominated to rapidly proliferate in order to remove the presence of the telltale pathogen in its entirety.
Omenn syndrome and severe combined immunodeficiency
With the overall health of the human body in mind, Omenn syndrome and Severe Combined Immunodeficiency (SCID) directly reveal the utterly destructive havoc caused by the absence of an operable VDJ mechanism. Moreover, due to the fact that both disorders involve RAG gene mutations, they further show that the entire production of variable antigen binding receptors is utterly helpless without RAG proteins.
Omenn syndrome, for instance, involves missense mutations in either of the RAG genes, which leads to a severe disruption in the RAG proteins’ joint function while still allowing some recombination to take place since one RAG allele is still normal. Thus, VDJ recombination activity is maintained partially, at a rather crippling rate. Characteristic symptoms of Omenn syndrome include a rash that either appears at birth or “a few days afterward,” severe loss of hair, eyebrows, and eyelashes due to the rash, and coexisting opportunistic infections. SCID, on the other hand, involves nonsense mutations in the RAG genes, leading to the complete loss of RAG protein function and thus of the VDJ mechanism. Infants born with such a disorder are susceptible to all infections, including common viral pathogens and opportunistic pathogenic diseases. The persistence of these infections consequently leads to “pneumonitis, enteritis, failure to thrive,” and, if left untreated, death occurs.
However, RAG gene deficient disorders, such as Omenn syndrome and SCID, still gravely reveal the severity of destruction caused to the human body if the VDJ mechanism is not working properly.
With all this information put into perspective, it is truly astonishing how VDJ recombination uses randomization to continually create the B and T cell receptors of great diversity and consequently keep the adaptive immunity strong against the ever potent pathogenic microorganisms. The role of VDJ recombination in the activation of genes in the generation of antibodies and T cell receptors diversity is enormous. VDJ recombination allows the expression of millions of unique combinations. With the identification of the RAG1 and RAG2 protein enzymes along with the others expressed in the nonhomologous end joining mechanism, it is clear the diversity possibility is endless. Despite the structure of the enzymes being a mystery, the diversifying function is known. The variable, diverse, and joining gene segments are solely responsible for encoding the diversity seen in immunoglobulins and T cell receptors. As studies continue to uncover more mechanisms in the primary lymphoid tissue, the more possibility for diversity there will be within the VDJ mechanism.
The affinity maturation, which is thought to be driven by somatic mutation, might also involve the VDJ recombination and receptor editing. Edited receptors would usually comprise the original heavy chain with an altered light chain.
Despite that we are developing a reasonable understanding of the factors that dictate the process of VDJ recombination at different TCR loci, there is still more that is not yet clear. It is still not clear how RAG proteins find RSSs and assemble synaptic complexes. How and to what sites is RAG recruited or are RSSs recruited to RAG.
[Crossref] [Googlescholar][Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
[Crossref] [Googlescholar] [Indexed]
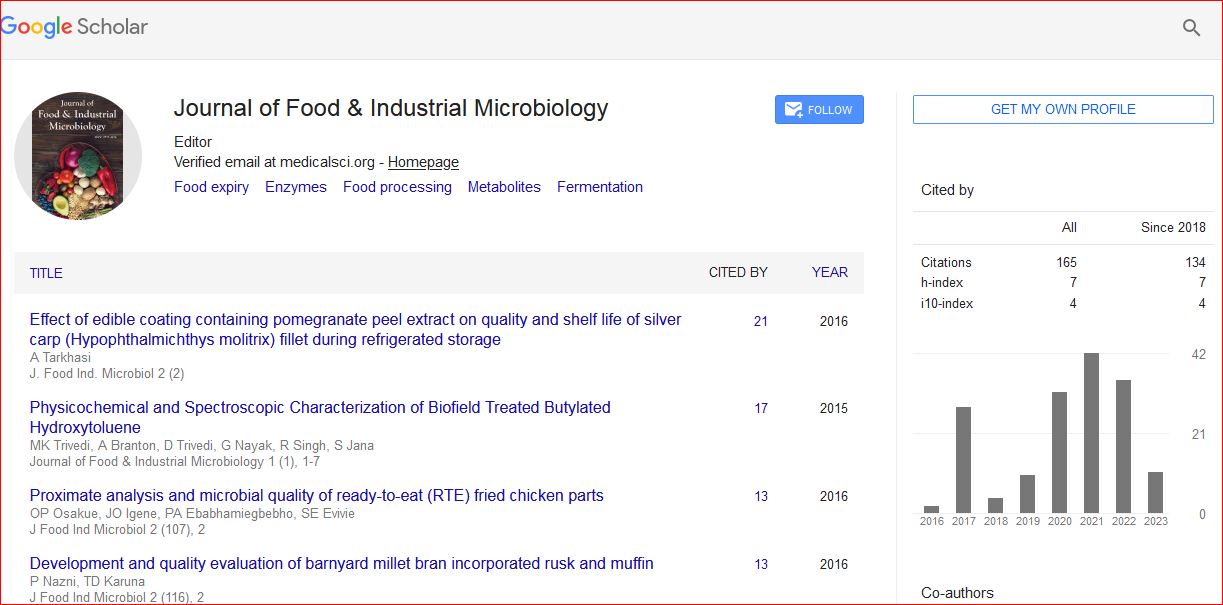
Journal of Food & Industrial Microbiology received 160 citations as per Google Scholar report