Short Communication - (2025) Volume 10, Issue 1
Received: 01-Mar-2025, Manuscript No. jib-25-168760;
Editor assigned: 03-Mar-2025, Pre QC No. P-168760;
Reviewed: 15-Mar-2025, QC No. Q-168760;
Revised: 20-Mar-2025, Manuscript No. R-168760;
Published:
27-Mar-2025
, DOI: 10.37421/2476-1966.2025.10.268
Citation: Kant, Casalone. “The Metabolic Landscape of Dendritic Cell Function and Differentiation.” J Immuno Biol 10 (2025): 268.
Copyright: © 2025 Kant C. This is an open-access article distributed under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
DC development and function rely on dynamic changes in cellular metabolism. During steady-state conditions, immature DCs maintain a basal metabolic rate largely reliant on mitochondrial OXPHOS and fatty acid oxidation. This quiescent state supports long-lived survival and antigen surveillance. Upon activation through Pathogen-Associated Molecular Patterns (PAMPs) detected by Toll-like Receptors (TLRs), DCs undergo a metabolic shift toward increased glycolysis, even in the presence of oxygen-a phenomenon known as aerobic glycolysis or the "Warburg effect." This reprogramming supports rapid energy needs, biosynthesis of macromolecules, and robust cytokine production. Classical DCs are further divided into cDC1 and cDC2 subsets. cDC1s, which excel at cross-presentation of exogenous antigens to CD8+ T cells, depend heavily on OXPHOS and fatty acid metabolism [2].
Disruption of mitochondrial metabolism, such as inhibition of FAO with etomoxir, shifts DCs toward an inflammatory phenotype, underscoring the immunomodulatory role of mitochondria. Furthermore, mitochondrial ROS (mtROS) generated during respiration can act as signaling molecules, activating inflammasomes (e.g., NLRP3) and promoting IL-1β secretion, thus linking metabolism with innate immunity. Fatty acids serve both as structural components of membranes and energy substrates through β-oxidation. In DCs, FAO supports OXPHOS and facilitates tolerogenic programming. Flt3L-induced DCs, particularly cDC1s, show a dependency on FAO during development. PPARγ and PGC-1α are transcriptional regulators that enhance FAO and mitochondrial function, contributing to DC subset specialization. Interestingly, lipid accumulation in DCs, often observed in the tumor microenvironment or in chronic infection, can impair antigen processing and presentation. Lipid-laden DCs show reduced cross-presentation capacity and an exhausted phenotype. This metabolic reprogramming, often driven by tumor-derived factors or hypoxia, represents a mechanism of immune evasion [3].
Amino acids such as glutamine, arginine, and tryptophan are critical for DC function. Glutamine provides carbon for the TCA cycle, supports nucleotide biosynthesis, and fuels OXPHOS in immature DCs. Upon activation, glutaminolysis shifts toward providing metabolic intermediates for cytokine production and epigenetic regulation. Arginine metabolism in DCs is catalyzed by nitric oxide synthase (NOS) and arginase, yielding nitric oxide (NO) and urea, respectively. NO can modulate T cell responses and induce apoptosis in neighboring cells, while arginase-mediated arginine depletion suppresses T cell proliferation, contributing to immune suppression in tumors and chronic inflammation [4,5].
Google Scholar Cross Ref Indexed at
Google Scholar Cross Ref Indexed at
Google Scholar Cross Ref Indexed at
Google Scholar Cross Ref Indexed at
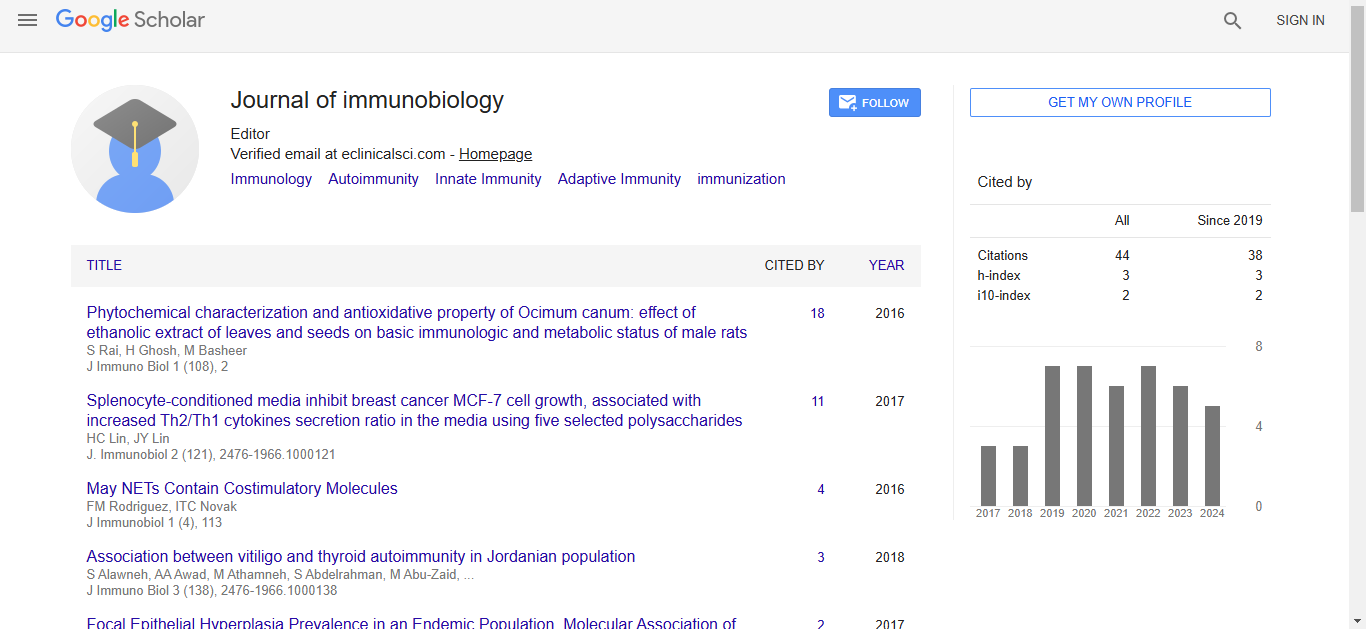
Journal of Immunobiology received 34 citations as per Google Scholar report