Research Article - (2021) Volume 6, Issue 4
Received: 07-Jun-2021
Published:
28-Jun-2021
, DOI: 10.37421/2476-1966.2021.6.163
Citation: El Amir, Azza, Nabil Wajeet, Eldaly Marwa M. and Hosney Mohamed. "Susceptible Role of Saccharin Mixture in Induction of
Inflammation through Action on Interleukin-10 and its Receptor" J Immuno Biol 6(2021): 163.
Copyright: © 2021 El Amir A, et al. This is an open-access article distributed under the terms of the creative commons attribution license which permits unrestricted
use, distribution and reproduction in any medium, provided the original author and source are credited.
Sodium cyclamate and saccharin mixture is a common artificial sweetener used as a food additive and it was found to be accompanied with increase relevance of inflammation including IBD. The causes of IBD are unknown, but many immune, genetic and environmental factors play a role as causative agents of that disease. The consumption of artificial sweeteners was related to the appearance of inflammation including IBD. The present study was aimed to evaluate the effect of sodium cyclamate and saccharin mixture as artificial sweeteners on immune system in albino mice through assessment of IL-10 cytokine and its receptor. Fifty male albino mice grouped into control group (n=20) receive only tap water and treated groups (n=30) received 60 mg/kg body weight sodium cyclamate and saccharin mixture in tab water for 12 weeks. Mice were sacrificed twice at the 6th and the 12th weeks. Collected blood measured low concentration of IL-10 cytokine by the sandwich ELISA. While, expression levels of IL-10R1 and IL-10R2 were assessed in colorectal tissue samples using Real time PCR. The results demonstrate significantly overexpression of IL-10R1 and IL-10R2 at 6th week than 12th week treated mice was recorded. Also, more congestion of RBCs, focal aggregation, focal necrosis, and focal infiltrations between glands in the lamina propria at 12th week treated group than 6th week treated group were observed. In conclusion, sodium cyclamate and saccharin mixture induced inflammation that may be risk for IBD by alteration in gene expressions of IL-10R1 and IL-10R2 in male albino mice.
Sodium cyclamate • Inflammatory bowel disease • Necrosis • Lamina propria • Focal necrosis
Artificial sweeteners are either synthetized chemical compounds or extracted from natural sources such as sucralose (Chlorination of sucrose). They are non-caloric, and have been approved as safe healthy substances by World Health Organization (WHO) and Food and Drug Administration (FDA) for usage by the people (Sardarodiyan and Hakimzadeh, 2016).
Saccharin is the oldest non-nutritive sweetener discovered in 1878 [1]. It was used as antiseptic agent in the treatment of urinary tract infections, later; it is used as sweetening flavor and then becomes a substituent for sugar in food and beverages [2]. Some studies on animals showed that saccharin may act as a promoter of carcinogenesis in the urinary bladder, so it was banned by the FDA in 1970 [3,4].
Cyclamate was discovered in 1937, FAO/WHO expert committee on food additives allows a maximum daily intake up to 11 mg/kg. It became more popular than saccharin as a beneficial solution for some health issues such as weight loss and decreasing blood sugar [5].
Qin [2] proved that the consumption of artificial sweeteners was related to an inflammation that may be at risk for different diseases including Inflammatory Bowel Disease (IBD). IBD are characterized by the usually features of inflammation such as damage of the intestinal epithelial tissue, lesions and crypt abscess as a result of inactivation of gut bacteria and defect in inhibition of digestive proteases associated with de-conjugated bilirubin [2,6].
The causes of IBD are unknown, but many immune, genetic and environmental factors play a role as causative agents of that disease [2]. The most important environmental factor that affects IBD is the changes in the composition of the intestinal microbiota that activate the over action of immune response and lead to the intestinal mucosal barrier dysfunction [2,6]. The epithelial monolayer protects the physical barrier by formation of a web of Tight Junctions (TJs). Structural changes or even altered expression in TJs proteins can lead to the development of inflammation. Furthermore, secretion of some pro-inflammatory cytokines such as tumor necrosis factorα (TNF-α) and interferon-É£ (IFN-É£) can increase TJ permeability and induce apoptosis of intestinal epithelial cells, that lead to mucosal inflammation [6].
Interleukin-10 (IL-10) is a soluble factor that is produced by type 2 helper T cells which inactivates the secretion of type 1 helper T cytokines including IL-2 and IFN-É£ [7]. IL-10 has pleiotropic effects on B cells, T cells, myeloid cells, and other cell types [8]. IL-10 is an immuno-modulatory, act as an anti-inflammatory cytokine and plays a vital role as ‘protective’ function in an inflammation including IBD [9]. IL-10 starts its activation when binds to its receptor; interleukin-10 receptor (IL-10 R).
IL-10R is a hetero-tetramer complex composed of two alpha molecules IL-10R1 (encoded by the IL-10 ra gene) and two beta molecules IL-10R2 (encoded by the IL-10 rb gene). IL-10R1 is expressed on immune cells, whereas IL-10R2 is expressed on somatic and immune cells. It was found that human patients with deleterious mutations in either IL-10 or its receptor develop severe inflammation including IBD. Also, mice with deficiency in either IL-10 or IL-10R can develop entero-colitis [10-12]. In mice, IL-10R signaling is important in T regulatory (Treg) cells to prevent development of colitis [13].
Therefore, the present study was aimed to evaluate the effect of sodium cyclamate and saccharin mixture as artificial sweeteners on the immune response of inflammation that may be at risk for IBD in albino mice through assessment expression of IL-10 cytokine and its receptor.
Ethics statement
The present study was approved by the Institutional Animal Care and Use Committee (CU-IACUC) of the Cairo University (CUIF3016).
Animal experiments and grouping
Fifty albino male mice (2-month age, weight 19-25 gm.) were supplied by Theodor Bilharz Research Institute (Giza, Egypt) and housed in Cairo University animal facility and provided a rodent diet and tape water under the following conditions: 25°C, 40%-70% humidity and 12:12 hour light/ dark. The artificial sugar pills are a mixture of 40 mg sodium cyclamate and 4 mg saccharin (Sussil, Germany).
Mice were randomly divided into control group (n=20) receive only tap water and experimental group (n=30) which received a mixture of sodium cyclamate and saccharin in tap water for 12 weeks with the dose (60 mg/kg body weight/day) [5]. Mice were euthanized as sacrifice twice at the 6th and the 12th week. Blood was collected from the jugular vein and kept overnight in the room temperature, then centrifuged, heat inactivated at 56°C water bath and concentration of IL-10 level was measured by the sandwich ELISA assay.
Small section samples of colon (2 cm), washed by cold phosphate buffered saline (PBS) and were applied to staining by Hematoxylin-Eosin (H&E) or immersed directly into RNA later solution for assessing gene expiration of IL-10 receptor using Real time PCR.
Enzyme-linked immunosorbent assay (ELISA)
Concentration of IL-10 in serum was determined using the ELISA kit (Due set ELISA; R&D systems, Minneapolis, MN) following the manufacturer's protocol.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
RNA was extracted from 16 frozen colorectal tissue samples from 6th week and 12th week control and treated mice (4 mice/ each group) using the Gene JET RNA Purification kit (Thermo Fisher Scientific, Inc., USA), according to the manufacturer's protocol and was stored at -80°C. A total of 1 μg total RNA was converted into cDNA using a high-capacity reverse transcriptase kit (Applied Biosystem, Inc., USA), according to the manufacturer’s protocol. Real-time PCR was performed using the Maxima SYBR-Green Master Mix kit (Thermo Fisher Scientific, Inc.) to amplify IL- 10 receptor 1 and 2 (IL-10R1 and IL-10R2) using Beta-Actin (β-Actin) as a housekeeping control gene. Primers used for qPCR were commercially synthesized by (Invitrogen, Inc., USA) and they are listed in Table 1. The reaction was detected with Applied Biosystem 7500 Step One Plus and reactions were done in triplicate. Each reaction was initially denatured at 95ÃÂ??C for 5 min, and then subjected to 40 cycles of the following: Denaturation at 95ÃÂ??C for 50 sec, and annealing and extension at 60ÃÂ??C for 1 min. Each sample was exposed to a final extension at 72ÃÂ??C for 10 min and finally held at 4ÃÂ??C. Melting curves were also conducted after amplification to ensure the reaction specificity. Following qPCR, Cq values were measured, ΔΔCq and fold expression were calculated to quantify the results [14].
| Gene | Direction | Sequence |
|---|---|---|
| IL-10R1 | Forward | 5'-AGGCAGAGGCAGCAGGCCCAGAATGCT-3' (Denning, 2000) |
| Reverse | 5'-TGGAGCCTGGCTAGCTGGTCACAGTAGGTC-3'(Denning, 2000) | |
| IL-10R2 | Forward | 5' GCCAGCTCTAGGAATGATTC3' (Denning, 2000) |
| Reverse | 5' AATGTTCTTCAAGGTCCAC-3'. (Denning, 2000) | |
| β – Actin | Forward | 5' -CTGTCCCTGTATGCCTCTG-3' |
| Reverse | 5' -ATGTCACGCACGATTTCC-3' |
Table 1: Primers sequences of target genes for RT-qPCR. IL-10R1: Interleukin 10 receptor 1 gene; IL-10R2: Interleukin 10 receptor 2 gene.
Histopathological study
Sections of colon (2 cm) were removed from all mice groups and washed by cold PBS, fixed after in 10% formalin for 24 hour, dehydrated in 70%, 80%, 90% and 99% alcohol and embedded in paraffin wax. 4 μm sections thickness were prepared by Rotary Microtome (Leitz) and de-paraffinized for staining with H&E staining. In addition, tissue sections were stained with alcian blue and Periodic Acid Schiff’s (PAS) staind. The combination of these two stains is more useful in differentiation between neutral and acidic mucins and also to study changes in the distribution of mucins in the tissues. Firstly sections stained with Alcian blue which stains all acidic mucins (Sialomucin and Sulphomucin) with deep blue stain followed by PAS stain that stains the neutral mucins with a brightly magenta color. The slides were covered by mounting media (Fisher Scientific) and examined using light microscope.
Measurements of IL-10 level in mice sera
The results of the present study demonstrate that there was a nonsignificant difference in concentration of IL-10 in serum between 6th week and 12th week treated group and compared to control group in male albino mice (PË?0.05; Figure 1). On the other hand, there was a significant decrease in IgA levels and significant increase in IgG levels between the two sacrifices in mice (P<0.05; Figure 1) (Table 2).

Figure 1: The levels of A) interlukin-10 (IL-10), B) IgA and C) IgG in serum of control mice and those treated with cyclamate-saccharine blend, after 6 and 12 weeks. Data is displayed as mean ± Standard Error of Mean (SEM). Bars marked with similar superscript letters are insignificantly different (P>0.05), whereas those with different ones are significantly different (P<0.05).
| Time (Weeks) | ||||
|---|---|---|---|---|
| 6 | 12 | |||
| Control | Treated | Control | Treated | |
| IL-10 | 42.00 ± 2.91a | 46.83 ± 7.11a | 47.60 ± 5.70a | 47.67 ± 5.88a |
| IgA | 28.17 ± 0.60b | 35.83 ± 1.68c | 25.50 ± 1.95b | 20.17 ± 0.60a |
| IgG | 257.67 ± 0.60a | 295.83 ± 24.23a | 255.17 ± 7.40a | 385.00 ± 10.09b |
Table 2: The levels of interlukin-10 (IL-10), immunoglobulin A (IgA) and immunoglobulin G (IgG) in serum of control mice and those treated with cyclamate- saccharine blend, after 6 and 12 weeks. Data is displayed as mean ± Standard Error of Mean (SEM). In the same row, means marked with similar superscript letters are insignificantly different (P>0.05), whereas those with different ones are significantly different (P<0.05).
Expression of IL-10R1 and IL-10R2 in colorectal tissues
The mRNA expression level of IL-10R1 and IL-10R2 genes from colorectal tissue samples at 6th week and 12th week treatment mice was assessed and changes were shown in Figure 2. Results showed that IL10-R1 and IL-10R2 levels were significantly overexpressed at 6th week treated mice than 12th week treated mice. Regarding to the levels of IL-10R1 increasing folds in colorectal tissues was 7.9-fold and 2.1-fold, respectively (P<0.001; Figure 2). While, the IL-10R2 increasing expression folds in colorectal tissues was 7.9-fold and 1.5-fold, respectively (P<0.001; Figure 2).

Figure 2: Comparison between fold expression levels of IL-10R1 and IL-10R2 mRNA in colorectal tissues at 6th week and 12th week treated groups and control group of mice using the Student’s t-test. Bars marked with similar superscript letters are insignificantly different (P>0.05), whereas those with different ones are significantly different (P<0.05).
Histopathological study
Sections from the control group showed normal histological structures; normal mucosal layer with lining epithelium and lamina propria of connective tissue with glandular structure and underlying mucosa, muscularis and serosa (Figure 3A).

Figure 3: Light photomicrographs of colorectal tissues from mice of control and treated groups stained with hematoxylin and eosin. a) Colorectal tissue of mice from control groups after 6 and 12 weeks. Histological examination showed average mucosa with average crypts (black arrow), average mucosal epithelial layer with average goblet cells (blue arrows), average muscularis mucosa (red arrow), average submucosa (green arrow), and average musculosa (yellow arrow) (X 400). b) Colorectal tissue sections from mice after 6 weeks of treatment showed moderate decrease in crypt numbers, moderate cryptitis and moderate reduction of goblet cells (black arrows), minimal congestion (red arrow) and intact mucosal epithelial layer (blue arrow) (X200). c) colorectal tissue sections from mice after 12 weeks of treatment showed atrophied ulcerated and edematous mucosa (black arrow) with atrophied crypts (blue arrow), markedly edematous muscularis mucosa (red arrow), edematous submucosa (green arrow), edematous musculosa (yellow arrow), focal inflammatory loci (thick red arrows), sever cryptitis (curved black arrow) and cryptic abscess (blue arrow heads) (X 200).
Sections of the 6th week treated group showed a slight manifestation in the colon such as oedema in the lamina propria between the glands in the mucosal layer, congestion of RBCs and slight infiltration of the inflammatory cells (leucocytes) in lamina propria of mucosal layer (Figure 3B). Sections of 12th week treated mice group, focal aggregations were recorded in the lamina propria of the mucosa in the mucosal epithelial layer of colon associated with desquamation of the lining mucosal epithelial layer and focal necrosis was also noticed in the mucosa as in (Figure 3C). Also, it was found focal infiltrations between the glands in the lamina propria of mucosa, necrotic epithelial cells, degeneration of crypts of liberckine (cryptic loss) and congestion of RBCs (Figure 3C).
Colonic walls in sections from the 6 and 12-weeks control groups showed normal average crypts with average positive blue mucoussecreting cells for acid mucins. Sections of 6-weeks treated group, colonic walls showed short crypts with minimal reduction of positive blue mucous- secreting cells for acid mucin. While; sections of 12-weeks treated group, the colonic walls showed a relatively average crypts with marked reduction of positive blue mucous-secreting cells for acid mucin (Figure 4).

Figure 4: Light photomicrographs of colorectal tissue sections of mice from control and treated groups stained histochemically using AB/PAS reaction to demonstrate different types of mucin. a) Colorectal tissue sections from mice of the control groups after 6 and 12 weeks revealed dominance of acid mucin-secreting goblet cells throughout the whole crypt length with increased concentration at the lowest two thirds of crypts and the goblet cells within the mucosal epithelial layer (black arrows) and the apical parts of crypts showed neutral mucin (red arrows) (X400). b) colorectal tissues section after 6 weeks of treatment showed short crypts with marked reduction of positive acid mucous-secreting goblet cells with specific localization at the lower part of crypts (black arrows) while the neutral mucin is evident at the upper two thirds of crypts and the through the mucosal epithelial layer (red arrows) (X 400). c) after 12 weeks of treatment, colorectal tissue sections showed relatively atrophied shorter crypts with severe reduction of positive blue acid mucous-secreting cells (black arrows), surface ulceration with prominent granular tissue was evident (red arrows) as well as cryptic abscess (thick black arrow) (X 400).
Sodium cyclamate and saccharin mixture are a common artificial sweetener used in the world as a food additive and its wide spread usage found to be accompanied with increase relevance of inflammation including IBD (Qin, 2012). Previous studies showed a counter effect of the artificial sweetener on gut microbiota, and colitis, which supported substantial evidence for the casual role of these artificial sweeteners in IBD [2]. On the other side, saccharin can also inactivate the bacterial growth, so it used as food preservative and an antiseptic [15].
The molecular reason of inflammation including IBD is largely unclear. Various genes are involved in the pathogenesis of inflammation including IBD; the similarity between these genes indicates the disruption of the adaptive and innate arms of the immune system [16-18], epithelial barrier function [19], autophagy process [20,21], and stimulation of the endoplasmic reticulum stress response [22] may cause susceptibility.
Despite the exact mechanism of inflammation including IBD is fully unclear, the interactions of genetic and environmental factors determinants have a vital role in the pathogenesis of disease [10,23-25]. Therefore, the present study was aimed to determine the effect of sodium cyclamate and saccharine mixture consumption on immune system of inflammation that may be risk for IBD in albino mice through assessment expression of IL-10 cytokine and its receptor.
It was found that patients with IBD have a deficient in either interleukin-10 (IL-10) or its receptor (IL-10R) or may suffer from a defect in the Janus kinase/Signal Transducer and Activator of Transcription (STAT) signaling pathway [6,10]. IL-10 is a key anti-inflammatory cytokine that is released predominantly by leukocytes such as B cells, T cells, monocytes, macrophages and dendritic cells, and also by some epithelial cells [13]. IL-10 works on both innate and adaptive immune cells and has large range of immuno regulatory activities that inhibits cytokine secretion, proliferation and co-activatory molecule expression of inflammatory immune cells [13]. IL-10 is anti-inflammatory pleiotropic cytokine that suppresses cytokines which initiate the inflammation cascade in the mucosal epithelial barrier. A vital role for IL-10 signaling in regulating intestinal mucosal homeostasis became appeared with the observation that IL-10-deficient mice promote spontaneous entero – colitis in human and mice [13].
The results of the present study demonstrate that there is a non- significant difference in concentration of IL-10 in serum between 6th week and 12th week treated group compared to control group in mice. The best explanation for that is the period of study was short and should be longer than 12 weeks.
The present study showed that IL10-R1 and IL-10R2 levels were significantly overexpressed at 6th week treated mice than 12th week treated mice, so, it was concluded that presence of more inflammation occurred at 12th week treated mice than 6th week treated mice. The best explanation for that is any defect in expression of IL-10 and its receptor can lead to inhibition of other pro-inflammatory cytokines, such as IFN-λ, IL-22, IL-26 resulting dramatic effects that ended by inflammation including IBD [10,13]. Also, there is another explanation deals with the signal transduction pathway (The receptor-associated Janus tyrosine kinases) that makes up after the assembly of receptor upon the IL-10 engaging. The inhibition of IL-10 and its receptors can lead to suppression in phosphorylation of STAT3 and STAT3- dependant genes. Disturbance in the transduction cascade or alteration in the gene expression of STAT3 can lead to inflammation including IBD [6,10]. A several mammalian cytokines regulate intracellular signaling by stimulating the mitogen-activated protein kinase (MAPK) pathway [26], as well as the Janus kinase/signal transducer and activator of transcription (STAT) pathway [27]. STATs are transcription factors that regulate cellular response via target gene expression, and STAT1, STAT3, and STAT4 have been found as IBD-associated susceptibility genes [10,28,29]. Stat3 plays a vital role in intestinal homeostasis and its preservative role to chemically induced colitis in mice [6,30].
The present histological results showed that after 6 week of sweetener mixture intake, changes in the colon tissues such as presence of oedema in the lamina propria between the glands, congestion of RBCs and slight infiltration of leucocytes in lamina propria of mucosal layer are observed. The best explanation for that is the absorption of artificial sweeteners in intestine can change the composition of intestinal microbiota [6] leads to induction of an overactive immune response that recruit the infiltration of leukocytes in the mucosal barrier. Alteration in the permeability and integrity of mucosal barrier will facilitate more infiltration of leukocytes and inflammatory cells. These observations became more illustrated in the colon sections after 12 week of sweetener mixture intake. In details, more congestion of RBCs, focal aggregation, focal necrosis, and focal infiltrations between glands in the lamina propria are observed. The best explanation for that is a huge increase in permeability of some pro-inflammatory cytokines, such as TNF-α and IFN-É£ in the mucosa that leads to increase the Tight Junctions (TJs) permeability in the mucosal barrier , apoptosis of intestinal epithelial barrier , epithelial cells necrosis, cryptic loss and ulcers [2]. TJs have an immune function including recognition of pathogen by production of mucus, so any defects in this production lead to more invasions of pathogen and intestinal particles into the mucosal barrier and induce inflammation.
In conclusion, the results of the present study demonstrate IL10-R1 and IL-10R2 levels were significantly overexpressed at 6th week treated mice than 12th week treated mice, so, it was concluded that presence of more inflammation occurred at 12th week treated mice than 6th week treated mice. Also, there is a non-significant difference in concentration of IL-10 in serum between 6th week and 12th week treated group compared to control group in mice. That may be due to the period of study was short and should be longer than 12 weeks. Furthermore, more congestion of RBCs, focal aggregation, focal necrosis, and focal infiltrations between glands in the lamina propria are observed at 12th week than 6th week treated group. Sodium cyclamate and saccharin mixture induced inflammation that may be risk for IBD by overexpression of IL-10R1 and IL-10R2 in male albino mice.
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
The authors declare that they have no conflicts of interest.
Experimental protocols and procedures were approved by Cairo University, Institutional Animal Care and Use Committee (CU-IACUC), CU/ I/F/30 /16 for animal use in research.
The data used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Azza El Amir, Wajeet Nabil and Mohamed Hosney have conceived, designed, planned, and supervised the research point. Marwa M. Eldaly, Wajeet Nabil and Mohamed Hosney performed the experimental work. Azza El Amir, Wajeet Nabil and Mohamed Hosney analyzed and interpreted the data. All authors participated in writing the manuscript, reviewed, and edited the final version of the manuscript. All authors approved to the final version of manuscript.
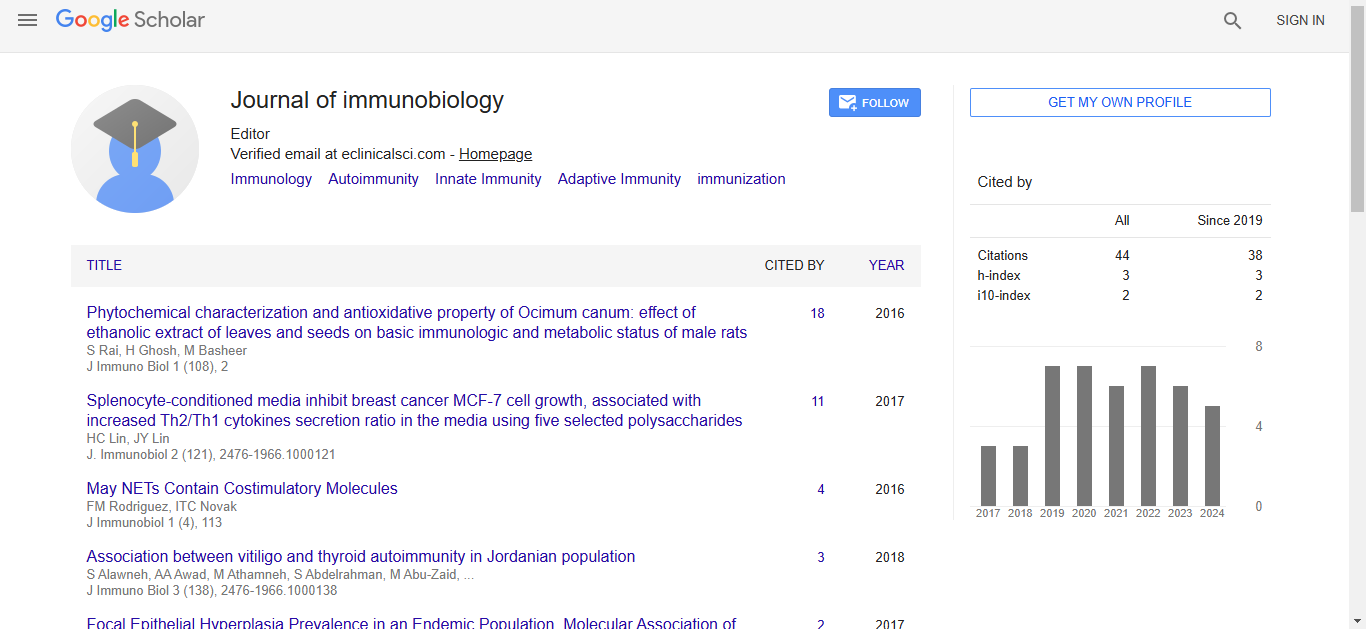
Journal of Immunobiology received 34 citations as per Google Scholar report