Brief Report - (2025) Volume 10, Issue 1
Received: 01-Mar-2025, Manuscript No. jib-25-168750;
Editor assigned: 03-Mar-2025, Pre QC No. P-168750;
Reviewed: 15-Mar-2025, QC No. Q-168750;
Revised: 20-Mar-2025, Manuscript No. R-168750;
Published:
27-Mar-2025
, DOI: 10.37421/2476-1966.2025.10.263
Citation: Campos, Noushmehr. “Epigenetic Regulation of Immune Cell Differentiation and Function.” J Immuno Biol 10 (2025): 263.
Copyright: © 2025 Campos N. This is an open-access article distributed under the terms of the Creative Commons Attribution License which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Epigenetic regulation refers to heritable and reversible modifications to chromatin structure that influence gene expression independently of DNA sequence changes. The major epigenetic mechanisms include DNA methylation, histone post-translational modifications, chromatin remodeling, and regulation by non-coding RNAs. These modifications collectively determine the accessibility of transcription factors to DNA, thus shaping transcriptional landscapes that define immune cell identity and functional states. DNA methylation, primarily occurring at cytosine residues within CpG dinucleotides, is a critical regulator of gene expression and lineage commitment in immune cells. Methylation typically represses gene transcription by impeding transcription factor binding or recruiting repressive protein complexes. During hematopoiesis, lineage-specific DNA methylation patterns guide the differentiation of multipotent stem cells into various immune cell types [2].
Traditionally, immunological memory has been attributed solely to adaptive immune cells. However, recent evidence reveals that innate immune cells such as monocytes and NK cells can also develop a form of memory-termed â??trained immunityâ?-through epigenetic reprogramming. Exposure to certain stimuli (e.g., β-glucan or BCG vaccine) leads to lasting changes in chromatin accessibility and histone modifications, resulting in enhanced responses to subsequent infections. Trained immunity is characterized by increased H3K4me3 and H3K27ac at promoters and enhancers of inflammatory genes, allowing for rapid and robust responses upon restimulation. In contrast, endotoxin tolerance-a phenomenon observed after repeated exposure to LPS-induces repressive marks like H3K9me3 and DNA hypermethylation at proinflammatory loci, thereby dampening responses. These opposing epigenetic outcomes illustrate the flexibility and context-dependence of innate immune training. Epigenetic aberrations contribute to the pathogenesis of numerous immune-related diseases [3].
The reversibility of epigenetic modifications presents a promising avenue for therapeutic intervention. Epigenetic drugs, including DNA methyltransferase inhibitors (e.g., azacitidine, decitabine) and HDAC inhibitors (e.g., vorinostat, romidepsin), are already in clinical use for hematological malignancies and are being investigated in autoimmunity and chronic inflammation. Epigenetic therapy can restore immune function by reactivating silenced genes, promoting T cell infiltration, or reprogramming suppressive myeloid cells in the TME. For instance, combining epigenetic drugs with immune checkpoint inhibitors has shown synergistic effects in preclinical cancer models by reversing T cell exhaustion and enhancing antigen presentation [4,5].
Google Scholar Cross Ref Indexed at
Google Scholar Cross Ref Indexed at
Google Scholar Cross Ref Indexed at
Google Scholar Cross Ref Indexed at
Google Scholar Cross Ref Indexed at
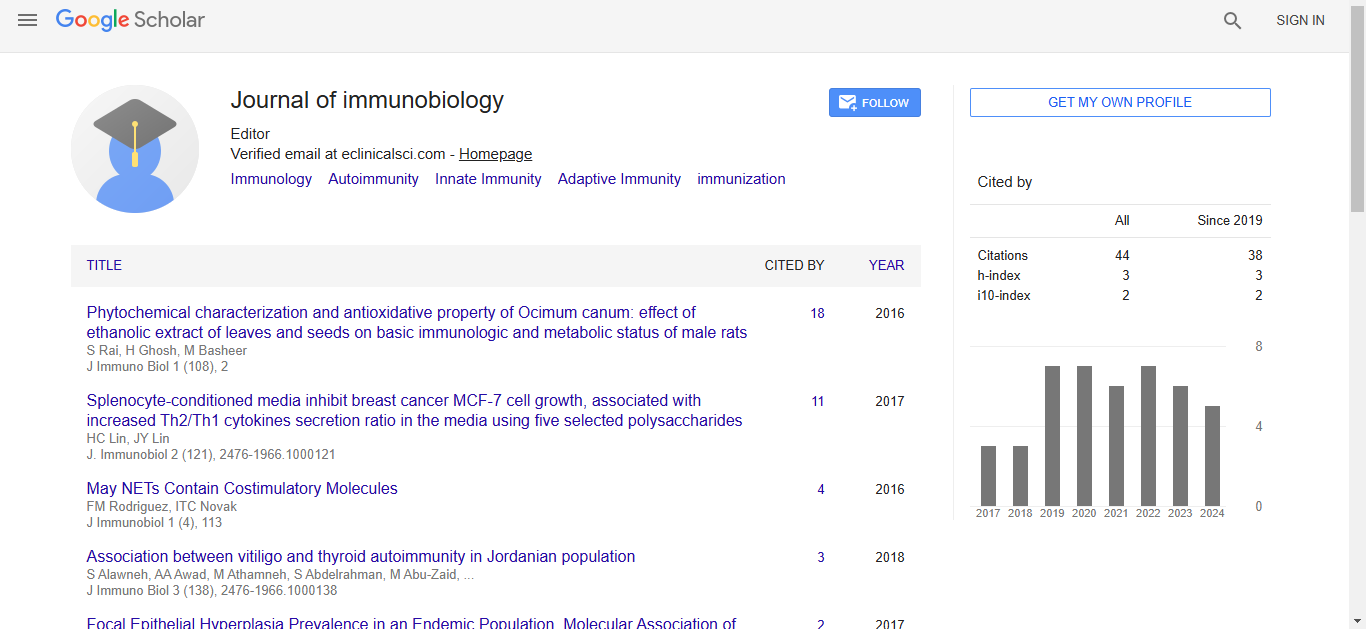
Journal of Immunobiology received 34 citations as per Google Scholar report