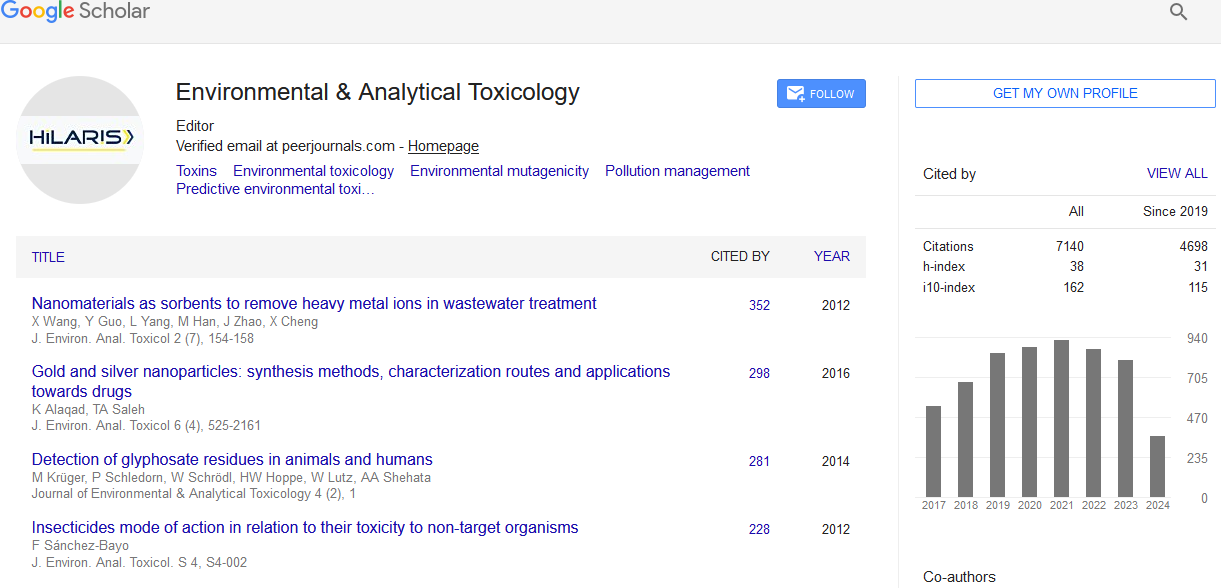
Review Article - (2022) Volume 12, Issue 3
Received: 24-Jan-2022, Manuscript No. JEAT-22-51677;
Editor assigned: 26-Jan-2022, Pre QC No. JEAT-22-51677;
Reviewed: 09-Feb-2022, QC No. JEAT-22-51677;
Revised: 24-Mar-2022, Manuscript No. JEAT-22-51677;
Published:
31-Mar-2022
, DOI: 10.37421/2161-0525.22.12.647
Citation: Saini Pradip Kumar, Prasad Shambhoo,and Bhati Jitender. "Transgenic Techniques for Iron, Zinc, and Iodine (IZI) Bio fortification in Crops." J Environ Anal Toxicol 12 (2022): 647.
Copyright: © 2022 Saini PK. This is an open-access article distributed under the terms of the creative commons attribution license which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
Micronutrient malnutrition will become more prevalent as the world's population grows, and it currently affects over three billion people globally [1]. In persons living in low-income countries, malnutrition deficiency, sometimes known as "hidden hunger," increases the risk of infectious illness and death from diarrhoea, measles, malaria, and pneumonia (WHO, 2000) [2]. Malnutrition has severe and long-lasting repercussions, which can even be passed down from generation to generation [3]. Malnutrition during pregnancy raises the chance of death and stunts growth, resulting in Low Birth Weight (LBW) and jeopardizing the child's life [4]. Malnutrition continues to be a serious public health issue in developing and impoverished countries [5]. People's diets are less diverse now than they were 30 years ago, resulting in micronutrient deficiencies, particularly iron (Fe), zinc (Zn), and iodine (I) [6]. Fe and Zn are critical trace elements for a range of metabolic functions [7]. Deficiencies in zinc and iron were ranked fifth and sixth, respectively, among the top ten risk factors for disease burden over the world [8]. Because of blood losses during menstruation and childbirth, micronutrient deficiencies are common in children and even more so in women [9]. Furthermore, in underdeveloped nations, micronutrient insufficiency is exaggerated by a lack of understanding and affordability of various and balanced meals, dietary patterns, and a high frequency of infectious diseases [10]. Various strategies such as food diversity, pharmacological supplementation, and fortification have been emphasized to fight nutritional deficit, particularly mineral inadequacy [11].
Current malnutrition-prevention strategies
The greatest strategy to prevent and even eliminate micronutrient deficiency is to eat a well-balanced diet rich in micronutrients [12]. Supplementation, dietary fortification, homestead food production, and crop bio fortification are all conventional but effective ways to manage malnutrition and micronutrient deficiencies [13]. The most practical technique of preventing vitamin deficit in people is to fortify diet with micronutrients [14]. Food fortification, as an approach for filling dietary gaps, has the added benefit of being able to give nutrients to vast portions of the population without necessitating drastic changes in eating habits [15]. Some countries have already begun to implement vitamin A fortification of butter, margarine, and sugar, as well as iodine fortification of salt, vitamin fortified milk, and vitamin B fortified cereals, all of which have a lengthy history [16]. Food fortification that is mandated rather than optional is more effective and has been used successfully for decades around the world [17]. Fortification of milk and oil is required in 14 and 27 nations, respectively. In 134 nations, salt fortification with iodine and fluoride is also required [18]. Around 83 countries are now using fortification legislation to supplement basic foods with micronutrients [19]. However, there are certain disadvantages to current food fortification procedures [20]. Food fortification is usually only possible in nations with well-developed, well-monitored, and well-regulated pharmaceutical and food processing industries [21]. Another significant disadvantage of supplementing and fortification systems is that they have recurring expenditures year after year, and their performance is contingent on funding [22]. This strategy works especially well in countries where the poorest people supplement their diets with small amounts of processed goods [23]. Unfortunately, consumption of industrially processed foods is small in the poorest developing countries, where the majority of the poor, particularly the farming population, rely on their own products for nourishment [24]. As a result, the reach of these food fortification programmers’ may be limited, particularly in developing countries' rural areas, where the majority of the poor live [25]. In India, obligatory fortification began in 1953 with the fortification of hydrogenated vegetable oil with vitamins A and D. To manage Goiter, salt fortification with iodization was used in 1998 [26]. West Bengal was the first to fortify wheat flour in 2000, followed by the Andaman and Nicobar Islands [27]. Micronutrient deficits are treated in industrialized nations by supplementation and fortification [28]. In poorer nations, however, extensive implementation of dietary diversification and food fortification/supplementation programmes is hampered by low income and poor market access for the target population [29]. Unfortunately, none of these measures based on socioeconomic development are effective in combating micronutrient deficiency [30]. Furthermore, the success of the aforementioned tactics necessitates societal behavioral changes that rely on literacy, communication, social marketing, and repeated investments [31]. Numerous investigators' global experience has confirmed that, rather than a single more expensive technique, a combination of several less expensive approaches is required to prevent nutritional shortage [32]. Plant scientists are concentrating their efforts to develop methods to apply fertilizers and/or use plant breeding strategies to increase the concentrations and/or bioavailability of mineral elements in the edible portion of crop plants to address the occurrence of micronutrient deficiencies in human populations [33]. Bio fortification methods are classified as 'agronomic' (fertilizer-based) and 'genetic' (breeding-based) [34]. Bio fortification is a method of increasing the micronutrient content of agricultural produce by targeting and modulating mineral nutrient movement pathways (root uptake, transport, remobilization, storage, and enhanced bioavailability), 'pulling' nutrients from the soil and 'pushing' them to economically important parts of the plant in bioavailable forms [35]. Agronomic bio fortification, traditional plant breeding, and genetic engineering are the three basic methodologies for bio fortifying food crops that have been used to date [36]. The main focus of agronomical methods is on optimizing mineral fertilizer application and/or improving mineral element solubilization and mobilization in the soil [37]. By lowering 'anti-nutrient' concentrations, the latter two techniques aim to generate and/or improve plant cultivars with greater micronutrient accumulation capability and increased levels of bioavailability [38]. To overcome the shortcomings of supplementation/fortification, bio fortification” (breeding for higher mineral and vitamin content) of staple foods is a potential, practical, and effective approach for supplying nutrient-dense food to rural populations [39]. This strategy can complement existing initiatives by providing a long-term and less expensive means of eliminating under-nourished populations that rely on supplementation and commercial fortification for nutrition [40]. The bio fortification strategy entails one-time fixed costs for creating breeding procedures, breeding nutritional quality features into present crop varieties, and adapting these varieties to a variety of conditions [41]. After nutritious varieties have been disseminated, this technique will require minimal recurrent investments [42]. Furthermore, the expenditures do not rise in proportion to the number of people, and the advantages may be distributed globally, particularly to underdeveloped countries [43]. Finally, there will be no yield penalty if you breed for higher trace mineral density in the consumed plant portions [44]. The two approaches to bio fortifying crops with minerals such as iron and zinc include conventional and molecular breeding, as well as genetic engineering techniques [45]. Because polygene’s with small effects influence the uptake and accumulation of micronutrients in edible portions of plants, traditional breeding-based bio fortification efforts have had only limited effectiveness [46]. Furthermore, the success of this method is largely dependent on the natural variance in the gene pool [47]. Genetic engineering will be a viable option for enhancing micronutrients at targeted levels if there is insufficient genetic variability and fixable major gene effects [48]. Despite the enormous efforts made through traditional plant breeding programmes, utilizing the field of genetics (quantitative genetics, heterosis, transgressive segregants, mutational breeding, marker assisted breeding, QTL mapping, and so on) to entrap natural genetic variations for micronutrients and vitamin accumulation, there is still a long way to go: the dream of nourishing the world's population [49]. This is primarily due to a number of flaws and threats to conventional breeding, including the need for sufficient genetic variations for a trait in the species, which may not be available for many economically important crops, the need for genes targeting the trait in sexually compatible plants, the long time period required for breeding to introduce a single as well as multiple traits (pyramiding traits) into locally adapted elite varieties without the risk of linkage drag, and the lack of genes targeting the trait in sexually compatible plants (e.g. cereal seeds, tubers etc.) as well as the soil's reliance on phyto-availability of mineral nutrients [50]. Furthermore, the inverse link between grain yield and grain mineral concentration has made it difficult for traditional breeding approaches to overcome such trade-offs [51]. Genetic engineering to make transgenic has also been used to transfer genes directly into elite genotypes as a modern weapon to combat mineral deficit [52]. Transgenic technologies are methods that can be used to improve genotypes by altering certain metabolic pathways [53]. These technologies provide the door for the modification of proteins, vitamins, carbs, lipids, minerals, and other metabolites, which will be discussed in the sections that follow [54]. However, two factors should ideally be considered while developing transgenics for nutrient bio fortification: (a) selection of a widely adapted genotype of an economically important crop, (b) nutrient accumulation in the edible section of the crop plant without compromising plant physiology, development, or economic yield [55]. Bio fortification based on conventional breeding is a well-accepted strategy for improving micronutrients in crops [56]. Using conventional breeding, a large range of crops has been targeted for fortification [57]. When there is insufficient genotypic variation for the desired attribute within the species (e.g. provitamin A in rice), or when the crop is not amenable to conventional breeding, genetic engineering techniques offer a viable alternative to traditional breeding tactics (e.g. banana) [58]. New gene editing techniques such as transcription activator-like effect or nucleases and CRISPR/Cas9, as well as greater availability of fully sequenced genomes in staple crops, have opened up new possibilities for this bio fortification strategy [59]. Transgenes can be used to redistribute micronutrients between tissues, improve the efficiency of biochemical pathways in edible tissues, reconstruct selected pathways (for example, in the field of system biology), increase micronutrient bioavailability by removing anti nutrients, and transfer multiple genes in a single plant [60]. As a result, bridging the gap between plant breeders and molecular biologists is critical for harnessing the power of genetic alteration for agricultural plant bio fortification [61].
Transgenic approaches for improvement of iron, zinc and iodine concentrations
Because minerals are not produced in plants, they must obtain Fe and Zn from the rhizosphere and immediate environment [62]. Various crops have been genetically modified to increase mineral content, particularly Fe and Zn [63]. Transgenic techniques to boost Fe and Zn content in crops have primarily focused on modulating transporter expression in order to improve plant uptake and utilization efficiency [64]. Phytic acid, for example, is an anti-nutritional component that should be reduced. In contrast to Fe and Zn, transgenic methods [65]. In the sections that follow, we'll go through some of the Fe and Zn genes and transgenic techniques used in important grains [66]. (See Table 1)
| Sl.No. | Crops | Nutrient | Gene/Protein |
|---|---|---|---|
| 1 | Rice | Fe and Zn | HvNAS1 |
| Fe and Zn | fumigates phytase | ||
| Fe | OsYSL2 | ||
| Fe | OsIDEF1 | ||
| Fe | Ferritin | ||
| Fe | Lactoferrin | ||
| Fe | Metallothionein-like protein | ||
| Fe | OsNAC5 | ||
| Zn | OsHMA1 | ||
| 2 | Wheat | Zn and Fe | OsNAS2 |
| Zn and Fe | phy A | ||
| Zn and Fe | phy A | ||
| Fe | TaVIT2 | ||
| Fe | GmFerritin | ||
| Zn | NAM-B1 | ||
| 3 | Soybean | Fe and Zn | Phytase |
| 4 | Barley | Fe | AtZIP1 |
| 5 | Maize | Fe | Ferritin and lactoferrin |
| 6 | Pea | Fe | brz and dgl |
| 7 | Tobacco | Fe | AtNAS1 |
| Tobacco | Iodine | HMT, S3H and SAMT | |
| 8 | Arabidopsis | Fe | AtNRAMP3 and AtNRAMP4 |
| Arabidopsis | Iodine | hNIS |
Table 1. Biofortified transgenic crops with increased iron, zinc, and iodine.
Iron (Fe)
Rice, wheat, and maize are targeted in large bio fortification programmes to address micronutrient inadequacies since they provide more than half of the caloric need globally [67]. Lactoferrin (a Fe-chelating glycoprotein) and ferritin were used in experiments to raise the Fe content of the endosperm [68]. Lactoferrin is abundant in human milk (1-2 g/l) (LF). Created transgenic rice grains with the human LF gene under the control of the rice glutelin-1 promoter to boost Fe content for future use in newborn formula [69]. Heterologous protein expression was much higher than control, reaching 0.5 percent of grain weight, and bioavailability was validated using a human Caco-2 bioassay [70]. The hLF gene was expressed in transgenic japonica rice and that it accounted for about 1.5 percent of total soluble protein [71]. Ferritin, a localised protein found in plant plastids, is a significant non-toxic Fe storage form that can release Fe as needed for metabolic processes [72]. Ferritin is a widespread protein that stores around 4,500 Fe atoms in an accessible state [73]. As a result, increasing Fe accumulation through ferritin gene expression controlled by endosperm-specific promoters is an essential method for Fe bio fortification [74]. Overexpression of ferritin in numerous crops enhanced Fe content and bioavailability, according to studies produced rice transformants of Soyfer H1 under the endosperm-specific GluB1 rice promoter to boost Fe accumulation in endosperm of brown rice seeds, and showed a threefold increase in grain Fe content compared to non-transformed lines [75]. Under the control of the maize ubiquitin promoter, soybean ferritin cDNA was also transferred in wheat and rice [76]. Due to the substantial role of leaves as a sink, the resulting transformants had higher Fe content in leaves than seeds [77]. That extra ferritin sequesters Fe in the leaves, decreasing Fe mobilization to the seeds, is also a possibility [78]. It used a strong endosperm specific globulin promoter to introduce soybean ferritin into rice, resulting in a 13-fold increase in ferritin protein expression [79]. The Fe content, on the other hand, had just a modest enchantment (30%). These findings revealed that, in addition to increased Fe storage, higher Fe transport from the soil and greater translocation within the plant system are required [80]. Genetic engineering proved to be a promising technique for Fe bio fortification in cereals in the rice trials [81]. In comparison to rice, however, very little work has been done to enhance the Fe content of wheat and maize grains, particularly in the endosperm [82]. Cloned and analyzed wheat ferritin genes (TaFer1-A) and demonstrated that ferritin over expression in the endosperm of wheat can enhance Fe concentration. The TaFer1-A gene was over expressed in the endosperm under the control of the HMW glutein 1DX5 promoter, resulting in a 50-85 percent increase in wheat grain content [83].
Iron-binding protein gene insertion
To generate Fe buildup in the seed that is ingested by humans, the first transgenic technique includes inserting an iron binding protein gene (lactoferrin) under the control of an endosperm specific promoter [84]. Plants or crops that store iron in their seeds are targeted for this purpose. Many attempts have been made to improve the grain iron content of rice, for example [84]. Rice is advantageous for a variety of reasons, including its low allergenicity and lack of harmful chemicals that interfere with gene expression [85]. Using a powerful endosperm specific promoter, researchers were able to successfully express human lactoferrin in de-husked rice. It resulted in a 120 percent increase in iron content, which was suitable for supplementing newborns; however, one molecule of lactoferrin binds to only two ferric ions, so it was still insufficient to fulfil the daily requirements of adults [86]. In wheat, soybean ferritin increased iron content by 1.5 and 1.9 times, respectively [87].
An iron-chelator gene was inserted
Nicotian Amine (NA) is an iron chelator that is important for Fe homeostasis and assimilation [88]. In rice (Table 1), the ferritin gene co-expressed with the Nicoti Anamine Synthase (NAS) gene results in a 6-fold rise in Fe content, which is higher than the single gene method [89]. In another study, expressing NAS, OsNAS3, resulted in a 7-fold increase in Fe content in transgenic rice seeds [90]. Anemic mice fed these transgenic rice seeds for four weeks recovered from iron deficiency and anaemia, whereas mice fed nontransgenic rice seeds showed no change. HvNAS1 over expressed by OsActin1 promoter/35S promoter in transgenic rice showed a 5-10 fold rise in endogenous NA levels in the shoots and seeds, resulting in a 3-fold larger increase in Fe content in T1 seeds. Similarly, transgenic tobacco with nicotianamine synthase (AtNAS1) overexpression had higher Fe levels. Bio fortified pearl millet has also been found to have a 2-fold increase in iron compared to most current wheat cultivars. An increase in iron absorption of 5-10% has been documented in roughly 35 million persons who eat bio fortified pearl millet [91].
Increased production of enhancers that improve Fe absorption
It's been proven that some foods can help with iron absorption. Vitamins including b-carotene, ascorbic acid, and α-tocopherol, as well as amino acids produced from proteins after digestion, fall into this category. Ascorbic acid and citric acid have been shown to decrease Fe to a ferrous form and increase small intestine absorption. As a result, transgenic techniques to over-express ascorbic acid in conjunction with ferritin can be applied. Fe absorption has also been demonstrated to benefit from higher cysteine concentration. Over expression of rice metallothionein-like protein resulted in a 10-fold increase in cysteine concentration in rice. It has 12 cysteine residues per mol of protein, identical to metallothionein. Golden rice with high levels of b-carotene has been found to have improved iron absorption.
Zinc (Zn)
Zn is a necessary element that also serves as a cofactor for over 300 enzymes and 1000 transcription factors. The natural variation of grain Zn content in cereals is modest. As a result, boosting the zinc content of cereal grains is critical for human nutrition and metabolism. Manipulation of the Zn content in cereal grains, on the other hand, may be more difficult than manipulating the Fe content found high connections between protein content, Fe, and Zn content in their study. Gpc-B1 (grain protein content b1) is a wheat quantitative trait locus linked to higher grain protein levels as well as higher Zn and Fe levels. After introducing the Gpc-B1 locus from the wild tetraploid wheat (Triticum turgidum ssp. Dicoccoides) into different recombinant chromosome substitution lines into cultivated wheat, an increase of 10–34 percent in grain Zn, Fe, Mn, and protein was observed, indicating the role of Gpc-B1 in the remobilization of protein, Zn, Fe, and Mn from the leaves to the grains. Over expression of genes involved in Zn translocation and mobilization resulted in improved Zn bioavailability without a yield penalty, which is an important strategy to improve grain Zn. Many cation transporters have been discovered in rice, but only a few have been studied in terms of substrate selectivity, expression pattern, and cellular localization. Members of the ZIP (ZRT, IRT-related protein) and CDF (Cation Diffusion Facilitator) families are the most common cation transporter families identified as being important in Zn uptake and translocation. In a thaliana root cells, the ZIP protein IRT1 plays an important role in Zn absorption. Introducing 35S enhancer elements to over-express NA synthase (NAS) resulted in 2-3 fold increases in Zn concentration in paddy. Similarly, polished rice grains from transgenic rice expressing the barley nicotianamine synthase gene HvNAS1 under the control of the rice actin1 promoter gathered 2–3 times more Zn. Thousands of IR64 and IR69428 transformants are created at IRRI using soybean or rice ferritin and rice nicotianamine synthase (NAS2) over expressed genetic constructs, and the Zn and Fe content in those lines has exceeded the goal level from field trials. As a result of the over expression of NAS genes, nicotianamine is a promising target for Zn bio fortification. Furthermore, bio fortifying grains using NAS alone or in conjunction with ferritin has a lot of promise in terms of addressing global human mineral deficiency. Sufficient research has been done in several crop species, including wheat; rice, maize, and barley, to better understand the Fe and Zn pathways in grain. Despite numerous obstacles such as the root-shoot barrier and grain filling, wheat researchers use rice-developed technologies and resources to boost Zn content in wheat grain, resulting in improved wheat lines.
NAS gene family over expression
Nicotianamine (NA) is a widespread chelator of transition metals (such as Zn and Fe) found in higher plants and is responsible for the transport of metal cations over short and long distances. The NA synthase (NAS) enzyme is involved in the production of NA by S-adenosylmethionine trimerization. Metal (Zn and Fe) profiles differentially control genes that encode for NAS in a range of plant species, including Arabidopsis, rice, maize, and barley. Over expression of exogenous or endogenous NAS genes is the most common method of increasing NA concentration in a plant using recombinant DNA technology. Exogenous HvNAS1 (barley NAS gene) over expression in tobacco and Arabidopsis has resulted in a several-fold rise in zinc, iron, and copper (Cu) concentrations in both plant species' seeds (Kim et al., 2005). In another study, over expression of HvNAS1 in rice resulted in a 15-fold rise in nicotianamine concentration (relative to wild type), as well as 1.5 and 2.5-fold increases in zinc and iron concentrations in polished rice grains. The rise in Fe and Zn concentration was caused by constitutive expression of AtNAS1 (Arabidopsis NAS gene) in combination with endosperm-specific production of ferritin. Furthermore, by expressing the OsNAS2 (endogenous NAS gene of rice) and Pv Ferritin (bean ferritin) genes in wheat grains, a significant level of Zn and Fe was attained. On the other hand, only a few researches have been published on the use of endogenous NAS to promote Planta NA expression for improved metal uptake in plants. Over expression of the endogenous NAS genes OsNAS1, OsNAS2, and OsNAS3 in rice resulted in significant increases in NA, Fe, and Zn concentrations in the endosperm of all three transgenic populations. In a different study, the OsNAS2 over expressing population had 20 and 2.7-fold higher NA and Zn concentrations than the wildtype population. Over expression of OsNAS3 resulted in a 9-fold increase in NA, a 2.2-fold increase in Zn, and a 2.6-fold increase in Fe concentration in polished rice grains.
NAC gene family over expression
Plant senescence is influenced by NAC transcription factors. Senescence is a synchronized process in which a whole plant or a portion of it remobilizes nutrients to younger tissues or seeds before dying. Senescence has been linked to increased Zn and Fe remobilization in studies. NAM-B1 (a member of the NAC transcription factors) has been discovered to play an important role in the early onset of senescence in wheat, which leads to a greater Zn concentration in grains. Reducing the amount of phytic acid in the body phytic acid (also known as phytate) is an inhibitor and antinutrient molecule that chelates minerals (Zn and Fe) and reduces their bioavailability, making it the most common cause of mineral deficiency in the globe. Because humans lack the intestinal phytase enzyme, phytic acid forms insoluble complexes with metal ions, particularly Zn and Fe, in the gastrointestinal system, which may not be absorbed or digested. By inhibiting their reabsorption into the body, phytate can form complexes with endogenously produced minerals, such as Zn. To counteract mineral deficiency, it is essential to reduce the amount of phytic acid in edible sections of staple crops to promote mineral absorption. Reduced phytate concentration in the diet has been linked to higher zinc absorption, according to research. Wheat transgenic lines had a 4-115 percent increase in bioavailable zinc in phytase.
Iodine
Iodine is required for the manufacture of the thyroid hormones triiodothyronine and thyroxine, making it a vital mineral for human health. Inadequate iodine intake affects more than 2 billion people worldwide. Triiodothyronine and thyroxine are two hormones that play an important role in metabolic control. Iodine shortage causes a reduction in the synthesis of these hormones, which leads to the growth of thyroid tissue, which is known as goiter. Goiter caused by iodine deficiency affects more than 187 million people worldwide. Furthermore, iodine deficiency during pregnancy may disrupt the offspring's neurodevelopment, whereas it affects somatic growth and cognitive skills during childhood. In comparison to other fortification techniques, adding iodine to table salt is a common strategy to prevent iodine deficiency. However, bio fortification of crops with iodine is a more promising strategy to combat mineral malnutrition because it is more sustainable and cost-effective. Several micronutrients, such as iron, zinc, and folate, have been successfully fortified into staple crops in recent decades. However, no reports of recombinant DNA-based bio fortification of iodine in crops have been published to yet. This is due to the physiology of iodine in plants being poorly understood. Iodine fluxes across the cell membrane of root cells via anion channels and probable H+/halides transporters, according to certain ideas. However, such transporters have yet to be found at the molecular level. Iodine volatilization from above-ground plant portions such as the root and leaf, on the other hand, is considerably well described in diverse species. Iodine volatilization is caused by the action of halide ion methyltransferase and halide/thiol methyltransferase. Transgenic Arabidopsis was the sole study that used genetic techniques to improve iodine concentration. The human sodium-iodide symporter (hNIS) gene of thyroid glands was over expressed under the CaMV 35S promoter in this study, resulting in increased iodine uptake. Furthermore, the HOL-1 gene, which encodes for the HMT enzyme (which causes iodine volatilization), was knocked out, and a significant reduction in volatilization was seen in transgenic plants compared to wild-type.
Micronutrients are crucial for human nutrition, particularly in the treatment of malnutrition in children and women. Bio fortification programmers rely heavily on micronutrients such as Fe, Zn and Iodine. Transgenic breeding is a strategic tool that can increase the content of these micronutrients in staple grains by several folds. Because genes for those desired qualities are available, it is conceivable to increase micronutrients in many staple crops. Trait-specific techniques have provided proof of concept for improving micronutrients through transgenic. It may be able to employ a combination of genes to improve micronutrients at the same time. After regulatory issues are resolved, transgenic crops will be widely planted to combat malnutrition. Furthermore, genome editing techniques for plant genes, such as CRISPR-Cas, ZFN, TALEN, and others, have recently showed significant promise in crop development. Crop bio fortification should also be made possible with genome editing techniques. Though genome editing-based bio fortification is still in its early stages, it should be widely used to accelerate bio fortification in cereals and horticulture crops, particularly vegetables. Bio fortified crop types with improved nutritional properties, on the other hand, must be tested in clinical studies for bioavailability and impact on end-user health.
Dr. Shambhoo Prasad Sir (Associate Professor), Department of Plant Molecular Biology & Genetic Engreening, Acharya Narendra Deva University of Agriculture and Technology, Kumarganj, Ayodhya (UP) India, for his supervision, Thanks a lot in writing this review article and for the suggestions made.
[Crossref][Google Scholar][Pubmed]
[Crossref][Google Scholar][Pubmed].
[Crossref][Google Scholar] [Pubmed].
[Crossref][Google Scholar][Pubmed].
[Crossref][Google Scholar] [Pubmed].
[Crossref][Google Scholar] [Pubmed]
[Crossref][Google Scholar][Pubmed].
Environmental & Analytical Toxicology received 6818 citations as per Google Scholar report