Mini Review - (2023) Volume 12, Issue 1
Received: 02-Jan-2023, Manuscript No. MBL-23-90783;
Editor assigned: 03-Jan-2023, Pre QC No. P-90783;
Reviewed: 16-Jan-2023, QC No. Q-90783;
Revised: 21-Jan-2023, Manuscript No. R-90783;
Published:
28-Jan-2023
, DOI: 10.37421/2168-9547.2023.12.367
Citation: Wells, Alexandria. “Myeloid Cells are an Essential Component of Numerous Inflammatory Processes.” Mol Bio 12 (2023): 367.
Copyright: © 2023 Wells A. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
In order to coordinate plant life and adaptation, unique membrane receptors that interpret both native and foreign cues have evolved. In order to be able to detect ligands of varying chemical compositions, this large family of receptor proteins have developed ectodomains that are very different from one another. Understanding these signaling systems mechanistically will help answer important questions in cell biology. The latest structurally characterized receptor–ligand interactions and regulatory mechanisms, in addition to novel receptor folds, will be the primary focus of this review. Prostaglandins (PGs) are bioactive lipids that have a place with the eicosanoid class of mixtures. Prostaglandins are known for regulating inflammation, vasodilation, and platelet aggregation, among other physiologic and pathologic functions. Some aspects of the outcome of immune responses, particularly T cell trafficking and diapedesis are influenced indirectly by these characteristics alone. However, PGs also have direct effects on immune system cells that are heavily dependent on the microenvironment during exposure.
Ectodomains • Plant membrane • Polypeptides • Brassinosteroid • T cells
Vegetation depends on the impression of numerous local and unfamiliar flagging prompts to arrange development and advancement, and to adjust and answer an always evolving climate. Plant membrane receptor kinases are able to interpret and transduce these extracellular signals across the membrane, initiating function-specific responses, and they perceive many of these signals at the cell surface. An intracellular kinase domain, a single membrane-spanning helix, and an extracellular domain that structurally diversifies to be able to perceive a variety of chemical ligands in a specific manner are all shared by this large family of receptor proteins. Small organic molecules, peptides amino acids, sugars, or lipids are examples of this diverse range of ligands. Plant structural biology has recently focused a lot on figuring out how specific intracellular responses are triggered by the physical interaction between receptors and their corresponding ligands. The ligand typically functions as a molecular glue in both homodimerization and heterodimerization receptor activation mechanisms. Upon ligand binding, Leucine Rich Repeat (LRR) RKs typically use heterodimerization with another LRR protein, known as a co-receptor.
The phosphocode of the cytoplasmic kinase domains determines the functional downstream response. Indeed, studies utilizing chimeric receptors with swapped kinase domains have shown that the receptor kinase is the one responsible for calling for downstream specificity. RK with other extracellular domain topologies have recently been structurally determined, revealing novel folds and opening up new avenues for receptor–ligand strategies and receptor modulation despite the fact that the LRR-RK family has been the most extensively studied to date, certainly due to their relative abundance in the genome. In a similar vein, the group of cysteine-rich polypeptides (CRPs) has gained a lot of ground under the microscope of structural biology thanks to the discovery of novel binding modes and promising activation mechanisms. The latest structurally characterized receptor–ligand interactions and regulatory mechanisms will be the focus of this review. Snippets from the section Modulation of ligand sensing and receptor activation by LRR-RKs Up until this point, the heterodimerization model of the LRR receptor activation mechanism has been described. In this model, the LRR ectodomain of the receptor recruits a shape complementary co-receptor to complete the binding pocket upon recognition of the ligand. The two kinase domains are brought into close proximity by ligandinduced binding of the receptor and co-receptor ectodomains at the cell surface, allowing for transphosphorylation and activation of the cytoplasmic signaling cascade [1].
However, a class of peptides known as "Recognition of Cysteine-rich Peptides by Receptor Kinases" mediate numerous aspects of plant physiology and development. Although the sequence and size of CRP family peptides range from amino acids, they all share a conserved cysteine-rich domain that is thought to form intramolecular disulfide bonds. Beyond the LRRs, one can anticipate significantly different binding modes to their cognate due to their larger size and potential folding ability: new folds, architectures, and potential novel receptor– ligand interactions are provided by new RK topologies, which offer insights into additional receptor families. The structures of the final sensors described. The topology of PDLP lacks the cytoplasmic kinase and includes two and a transmembrane domain. They are connected to plasmodesmata and are thought to be involved in immune response and systemic signaling. However, this review has covered the most recent mechanistic advances in receptor ligand perception and regulation of plant RKs as well as their biochemical function. Perspectives and conclusions RK multimerization as a result of the ligand acting as molecular glue is necessary for the receptor activation mechanisms. The primary premise of one more layer of LRR-RKs flagging balance has been as of late uncovered for insusceptibility and brassinosteroid [2].
The enzymatic breakdown of arachidonic acid by phospholipase on membrane phospholipids is the source of all prostaglandins. Cyclooxygenases that are either inducible or constitutively the primary substrate for the production is produced by this reaction. By inhibiting the activity of COX enzymes, nonsteroidal anti-inflammatory drugs (NSAIDs), which are commonly used to treat pain, inflammation, and fever, reduce the formation of PGs. PGs also play a significant role in neoplastic disorders, as demonstrated by the fact that taking NSAIDs can lower the risk of developing colorectal cancer and even cause some adenomas to shrink. However, this aspect of their biology has already been extensively discussed and the focus of this review will be on how prostaglandins affect T cell function and phenotype, particularly in autoimmune (AI) conditions. Enzymes that are expressed in a manner that is specific to each cell and tissue produce and break down particular PGs. The enzymes cytosolic PGE synthase (cPGES), microsomal prostaglandin E synthase and inducible mPGES-1 are in charge of PGE2 synthesis from PGH2, while 15-hydroxyprostaglandin dehydrogenase (15- PGDH) is primarily in charge of PGE2 degradation. During inflammatory and protumorigenic responses, dysregulation of PGE2 biosynthesis has been extensively reported. Furthermore, PGE2 is present in high concentrations in many infectious agent-triggered pathophysiological conditions of the kidney and lungs [3].
In addition, when the proper resolution of inflammation is compromised, such as in chronic inflammatory conditions PGE2 levels rise and persist. The majority of the available PGI2 is produced by prostacyclin synthase (PGIS), a cytochrome P450 synthase that is found on the ER. H-Prostaglandin D synthase (PGDS), which is expressed in hematopoietic cells, is primarily responsible for the biosynthesis of PGD2. Lipocalin-Like Prostaglandin D Synthase (L-PGDS), which is highly expressed in the brain but can be found in adipose tissue, skin, and lymph nodes plays a smaller role in PGD2 biosynthesis than H-PGDS. Lastly, the PGF synthase but its function in T cells is unknown. In homeostatic conditions and during inflammation, the tissue-specific localization and concentration of PGs are controlled by the aforementioned enzymes and specific membranebound transporters. Figure depicts a summary of PG metabolism, receptors, and pharmaceuticals that target this pathway. It is essential to keep in mind that the level of complexity of PG metabolism, which includes a degree of redundancy in the availability of PGs metabolic intermediary molecules, is significantly higher than the overview that we provide here. We believe that it is beyond the scope of this review to provide a comprehensive biochemical description of all PG metabolism; others have already done so. Figure provides an overview of PG metabolism and sensing [4].
The PGE2 anti-inflammatory and immunosuppressive paradigm has evolved to include the realization that PGE2 can exhibit strong anti- and pro-inflammatory properties, and that the net effect of PGE2 is highly context-dependent, particularly in relation to the type and extent of PGE2 receptors expressed at a given time in a particular tissue. On the other hand, PGD2 and PGF2 also increase during inflammation before immune cells are recruited, but T cells have mostly found that they have anti-inflammatory effects. PGD2 is able to regulate CD4+ memory T cell trafficking across blood vascular endothelium which may constitute a PG-coordinated control level for T cell trafficking. PGI2 is highly secreted primarily by endothelial cells. The ability of human DC cells that have differentiated in the presence of PGD2 to stimulate T cells and encourage the differentiation of naive T cells into Th2 cells is also impaired. However, very little is known about the role that PGI2 plays in altering APC function. It is important to note that, depending on the inflammatory stimulus and the context all PGs can either promote or inhibit inflammation. T cell-expressed prostaglandin receptors are highlighted in purple, while constitutive enzymes are highlighted in grey and inducible enzymes are highlighted in red. The blue boxes indicate pharmaceutical compounds that target various enzymes or receptors [5].
T cells respond strongly to prostaglandin E receptors (EP) but contribute significantly less to PG pools overall. When considering how PGs affect myeloid cells, it is well known that PGE2 can both contribute to the resolution of inflammation and promote it. This is demonstrated by the fact that PGE2 can both suppress neutrophil and macrophage functions during tissue repair and mucosal barrier infection as well as in cancer and promote the influx and activation of neutrophils, macrophages, and mast cells. PGE2 can also enhance the antigen-presenting cell (APC) function of nearby T cells by acting as an enhancer. PGE2 induces the strong release of IL-12 and IL-23 in DCs with concomitant upregulation of CCR7 expression. However, PGE2 demonstrates many nuances in its both pro- and anti-inflammatory effects on DCs. Dendritic cells are frequently the most potent context-dependent APCs. Therefore, it is essential to take into consideration the effect that a particular PG profile can have on APC-T cell communications and their outcomes, as well as other cytokines, chemokines, and cell-cell signals.
None.
None.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
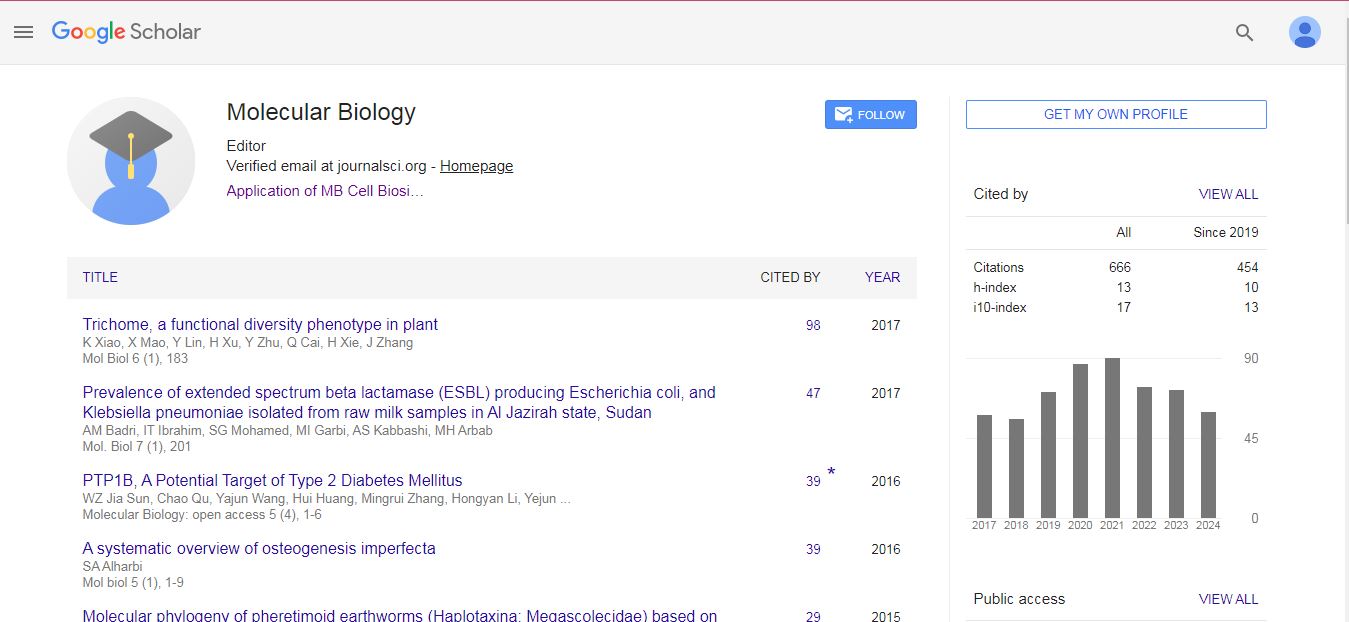
Molecular Biology: Open Access received 607 citations as per Google Scholar report