Commentary - (2022) Volume 6, Issue 3
Received: 02-May-2022, Manuscript No. JMBP-22-69809;
Editor assigned: 03-May-2022, Pre QC No. P-69809;
Reviewed: 16-May-2022, QC No. Q-69809;
Revised: 20-May-2022, Manuscript No. R-69809;
Published:
27-May-2022
, DOI: 10.37421/2952-8119.2022.6.152
Citation: Sanath, Shushma. “A Commentary on the Stage of Colon Carcinogenesis is Enter via Fungi.” J Microbiol Pathol 6 (2022): 152.
Copyright: © 2022 Sanath S. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
It has become increasingly clear that the intestinal microbiome is crucial for tissue homeostasis as well as the aetiology and treatment of a variety of inflammatory, immunological, and neoplastic illnesses. However, the majority of research to far have emphasised the function of microorganisms. Two publications in this issue of Immunity present persuasive evidence that the mycobiota, a different class of commensals, may also play a significant role in the development of colitis-associated colorectal cancer [1].
The caspase recruitment domain family, which includes the adaptor protein CARD9, has been shown to be crucial for the anti-fungal response in both mice and people. Card9-deficient mice are more prone to a variety of fungi, including Candida albicans, Aspergillus fumigatus, and Cryptococcus neoformans infections. Most Candida infections in people are caused by CARD9 deficiency, although an ascomycete infection has been identified in a recent investigation [2]. The C-type lectin receptors Mincle, Dectin-1, Dectin-2, and Dectin-3 are able to identify the fungus' b-glucans and a-mannans. The CARD9-MALT-BCL10 complex is assembled in response to ligand binding and the recruitment of the kinase Syk to the receptor. This combination then causes the NFkB and MAPK signalling pathways to be activated, influencing the production of cytokines.
In the two brand-new researches in this issue, CARD9's role in the AOM-DSS (azoxymethane and dextran sodium sulphate) model of CAC is investigated. Although Card9 loss increases intestinal tumour load in both situations, the postulated underlying mechanisms and the role of fungus appear to be distinct. Following the discovery that CARD9 expression was elevated in human CRC samples, it was decided to assess CARD9 performance in the AOM-DSS model [3,4].
Antifungal defenses
When spores of sexual or asexual (conidia) origin stick to the cuticle or are consumed with food, insects become infected by fungi. Hydrophobic coatings of fungus conidia encourage adhesion to insect exoskeletons, which promotes germination. As part of this process, germ tubes and appressoria, invasive structures that produce enzymes to break down proteins, chitin, and lipids, are formed. By triggering the prophenoloxidase cascade, the breakdown of cuticle proteins triggers a first-line defence response in the host, which eventually results in the production of melanin [4].
The injection of fungal cells into G. mellonella induces the expression of a broad spectrum of immunity-related effector molecules including numerous proteins with direct antifungal activity or the ability to inhibit virulenceassociated proteinases produced by Beauveria and Metarhizium species [5].
Avoidance or suppression
Entomopathogenic fungi have evolved a number of ways to reduce or prevent the antifungal immune responses in their insect hosts as an adaptation to their parasitic lifestyle. By generating so-called protoplasts or hyphal structures, which, in contrast to conidia and hyphae, lack a completely established cell wall, such fungi are able to circumvent immunological reactions in the hemolymph of the infected host. When penetrating hyphae enter the hemocoel shortly after infection, immunological competent hemocytes circulating in the hemocoel, such as plasmatocytes, phagocytose but do not destroy these hyphal entities [6].
Fungal responses against immunity
The reasons and effects of individual gene expression are unclear, but the abundance of genes linked to either fungal virulence or host resistance may encode proteins that interact with one another. As mentioned above, antifungal defence mechanisms in insects have been thoroughly researched, showing sensors that detect fungi that are invading as well as various immunity-related effector chemicals such antifungal peptides and inhibitors of fungal proteases [7].
There is growing evidence that parasitic fungi and the insects they parasitize have coevolved, leading to mutual adaptations. This shows that entomopathogenic fungi may recognize and block host immune responses.
No conflict of interest by author.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
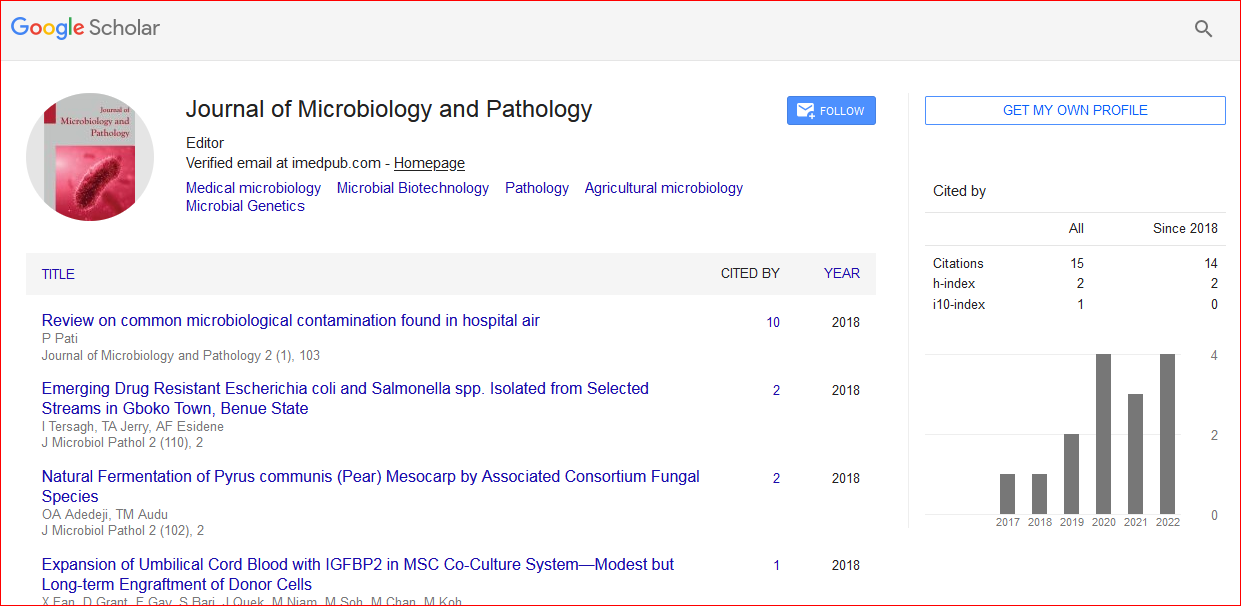
Journal of Microbiology and Pathology received 18 citations as per Google Scholar report