Serge Picaud
Abstract
Taurine is a free amino-sulfonic acid mostly provided by nutrition, which is present in huge amounts in the central nervous system. In retina where it signifies nearly half of the free amino-acid content Although more than 30 years ago. taurine reduction was found to trigger photoreceptor degenerat in the cats fed with a taurine-free diet. the mechanism of this taurine want still remains enigmatic. The effect of taurine on photoreceptor survival was the next confirmed in monkeys and in rats by administering an inhibitor substrate of the taurine transporter (Tau-T) and finally realised in mice by knocking out the taurine transporter. Taurine also looked to play a major role in photoreceptor development. Recently was established that the retinal toxicity of the antiepileptic drug vigabatrin, is caused by taurine depletion. Vigabatrin-treated rats and mice were found to exhibit lower plasmatic taurine absorption and taurine supplementation reduced the observed photoreceptor degeneration and the consecutive disorganization of the photoreceptor layers. However vigabatrin treated patients, the visual contraction is not only attributed to the photoreceptor degeneration but also to the retinal tumour cell (RGC) loss the RGC layer even appears as the main site of retinal damage.
Taurine Improves the Survival of Purified RGCs in Culture
To determine if taurine can affect directly to the RGC survival, it was applied on pure adult rat RGCs in the culture medium. The cell spotlessness of adult RGC culture was first evaluated by immunolabeling the RGC culture with two specific markers.
That most cells were immunolabeled with RGC markers NF-200 whereas a very few cells are positive for macrophage marker. taurine is a strong antioxidant and an osmotic regulation particle that requires Na+-dependent approval use to its cellular activities, we are inspected whether RGCs express the taurine transporter (Tau-T). RGCs were purified and used directly after the purification step (no culture) to measure the Tau-T expression level in newly purified RGCs. These data specify that RGC could generate the taurine uptake both in vitro and in vivo. GuanidinoEthane Sulfonate in RGC cultures. Interestingly, addition of GES with taurine significantly inverted the protective effect is used by taurine (+59%) on pure RGC cultures.
Taurine Reduced RGC Death in NMDA-treated Retinal Explant
Taurine was previously shown to prevent the glutamate excitotoxicity in neurons. We therefore discovered if taurine could also defend RGCs from glutamate excitotoxicity. Taurine can therefore partially avoid RGC losses from glutamate excitotoxicity. To determine the taurine could affect RGC existence in vivo, it was supplemented to the drinking water of DBA/2J mouse, a legalized genetic model of pigmentary glaucoma. Indeed, old animals were showing an increase in intraocular pressure (IOP) compared to C57BL/6J mice, taken as control. No clear modification of Tau-T immunolabeling was observed in DBA/2J mice as compared to C57BL6/2J in the ganglion cell layer (data not shown). These results show that taurine supplementation partly prevented to the RGC degeneration due to an elevated IOP in DBA2/J mice.
Discussion
Our previous experiments on vigabatrin-treated animals had suggested that taurine reduction can cause RGC degeneration. Here, we provide evidence to the taurine that can also promote the survival of adult RGCs in a pure and Clean culture. This result demonstrates a direct neuro protective action of taurine on RGCs, which is consistent with the taurine-elicited and resistance to the hypoxia in an preserved RGC cell line. The specific expression of the taurine transported in freshly purified rat RGCs (not cultured) advises that an intracellular mechanism they involved in taurine protective action. This theory was validated here by the loss of taurine neuro protection found in presence of taurine carrier inhibitor in GES. As a reported in the immortalized RGC cell line taurine neuro defence of RGCs could happen by reducing the intracellular calcium levels by preventing the opening of mitochondrial perviousness transition pores. Taurine was also shown to be needed for the translation of mitochondrial DNA.
In all our animal tests taurine supplementation increased by the two fold the plasma taurine concentration among although the starting concentration was different in mice and rats. upcoming studies will have to explore how the retinal taurine concentration is related to both this the transporter expression and circulating plasma concentration level in retinal endothelial cells. In humans, taurine supplementation has already been considered for the treatment of the various diseases and no human toxicity or undesirable side effects have been reported even at doses as high as 6 g/day. In fact, low doses (1.5 g/day) were already start to increase more than 2-fold the taurine plasma absorption in human subjects.
Conclusion
RGCs were isolated from retinae in adult Long-Evans rats (8-week old) with an immune criticizing technique, according to the protocols before described in young rats and adult animals. RNA was take out from freshly purified RGCs (not cultured), liver or kidney, full retina of Long-leaving rats, using the RN easy plus mini kit (Qiagen, Courtaboeuf, France). Reverse transcription were then made using the Super Script II (Invitrogen) incubation firstly at room temperature for 10 min, then at 42°C for 1 hour and finally at 70°C for 15 min in presence of Random Hexamer. Taurine was directed per by its addition into the drinking water for 4 months from 8- to 12–month old in DBA/2J mice, a genetic animal model for pigmentary glaucoma.
Immuno staining was performed on cryosections mice or rats eye cups by means of using anti-Brn-3a monoclonal antibody. RGC quantification depend on the POU4F1 (Brn3a) immune labeling because strong association was reported between POU4F1-immunopositive RGCs and retrogradely dye-stained RGCs, as early as 3 weeks and after the initial RGC lesion. In addition, POU4F1-immuno positive RGCs signify the majority of the RGC population (more than 75%) POU4F1/DAPI-positive RGCs were calculated from the whole vertical sections scanned with Digital Pathology System Nanozoomer (Hamamatsu, Massy, France) by 2 independent investigators unaware of the experimental groups. To evaluate the RGC thickness, the RCG numbers was normalized to the length of the retinal section measured with NDP-view software (Hamamatsu).
Note: This work is partly presented at 9th International Congress on Nutrition & Health February 20-21, 2017 Berlin, Germany
Share this article
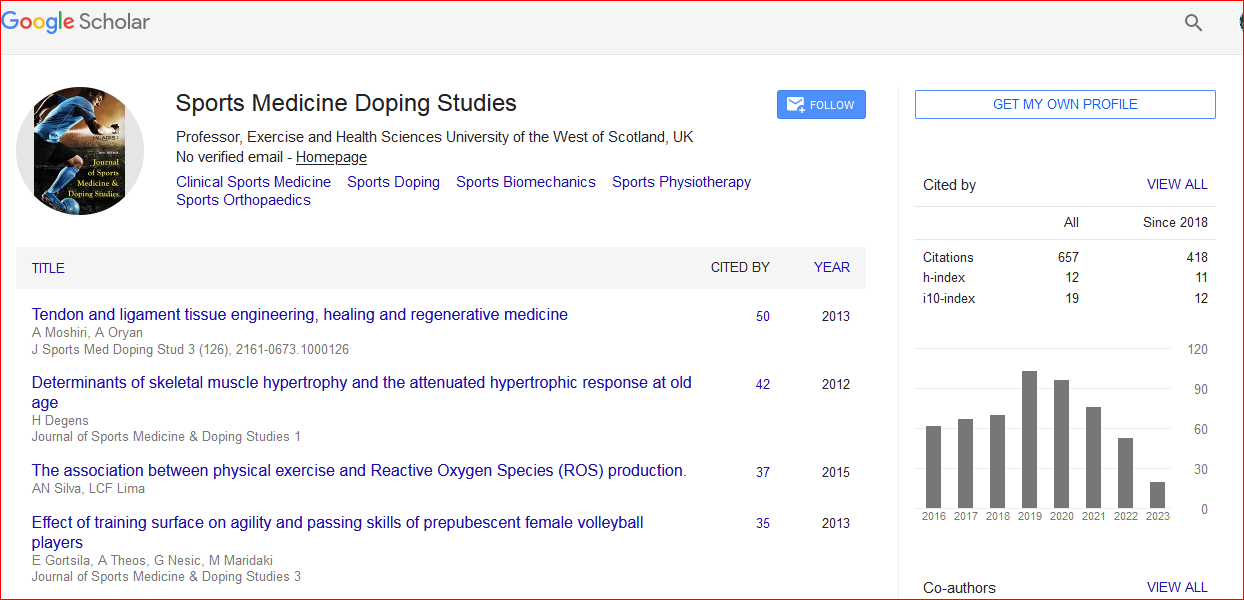
Journal of Sports Medicine & Doping Studies received 1022 citations as per Google Scholar report