Research Article - (2021) Volume 5, Issue 6
Received: 12-Nov-2021
Published:
03-Dec-2021
Citation: Souris, Marc, Léon Tshilolo, Destin Mbongi and Baltazar Phoba et al. “Suspected SARS-Cov-2 Original Antigenic Sin among Central and West African Populations.” Virol Curr Res (2021) 5: 144.
Copyright: © 2021 Souris M, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted
use, distribution and reproduction in any medium, provided the original author and source are credited.
More than a year after the emergence of COVID-19, significant regional differences in terms of morbidity persist, showing in particular lower incidence rates in some regions of Africa or Asia. The work reported here aims to test for a pre-pandemic natural immunity among populations in central and western Africa and a suspected SARS-CoV-2 original antigenic sin. To identify such pre-existing immunity, sera samples collected before the emergence of COVID-19 were tested to detect the presence of IgG antibodies reacting against SARS-CoV-2 proteins of major significance. Sera samples from blood donors of France also collected before the pandemic were used as control. The results showed a statistically highly significant difference for antibodies prevalence between the samples collected in Africa and the control samples. Our results suggest that in the tested African sub-regions the populations have been potentially pre-exposed before the COVID-19 pandemic to the antigens of a SARS-CoV-2-like virus.
Almost two years after the emergence of COVID-19, significant regional differences persist, showing the lowest incidence rates in sub-Saharan Africa, Southeast Asia, and Oceania. This trend was observed at the onset of the epidemic and has been confirmed during subsequent epidemic waves (Figure 1) [1].

Figure 1. COVID-19 incidence in the world from January 2020 to November 2021
(WHO). Note: 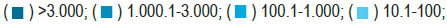

Several hypotheses have been proposed to explain this situation, including among others: the morbidity and mortality counts likely to be underestimated in some low- and middle-income countries due to limited epidemiological surveillance and/or public health screening activity; the population of sub-Saharan Africa is younger with only 2.3% of the population over 65 years old, whereas people over 65 years old account for more than three-quarters of the deaths related to COVID-19 in Europe (where this population represents more than 20% of the population); more rural living conditions may increase social distancing and reduce the spread of the disease; climatic and environmental conditions unfavourable to the virus and its spread; a natural immunity innate (nonspecific) or secondary due to previous contact with a coronavirus closely related to SARS-CoV-2 and sharing common antigenic profiles, and a suspected SARS-CoV-2 original antigenic sin (OAS).
The objective of this present study was to identify the later hypothesis of a pre-existing natural humoral anti-SARS-CoV-2 immunity among African populations by testing sera samples from repository collected several months before the COVID-19 epidemic started. We tested the presence of reacting antibodies against five SARS-CoV-2 proteins playing an essential role in virus attachment, fusion, entry and transmission. We present here our first results, obtained from 1,655 sera samples collected before January 2020 from people living in Democratic Republic of Congo (DRC), Cameroon, Republic of Congo (ROC), and Senegal.
SARS-CoV-2 virus pertains to the Betacoronavirus genus, which includes numerous virus species of Chiropterans. Chiropteran species are also hosts of several coronaviruses’ species close to the original strain of SARS-CoV-2 isolated in China [2]. SARS-CoV-1 and MERS-CoV viruses are also monophyletically placed with chiropteran coronavirus parental species [3]. In Cambodia and Myanmar, viruses closely related to SARSCoV- 2 were isolated from bat samples (Rhinolophus Shameli) collected before 2020 [4-6]. Globally, it can be observed that the spatial distribution of bats [7,8] matches the spatial distribution of countries with lower symptomatic circulation of COVID-19, especially in rural populations where the probability of contact with chiropterans is higher: ecology and behavior of bats, especially fruit bats (e.g.: mass frequentation of fruit orchards, roosting trees close to dwelling) may have favored the direct or indirect contact with humans. Such type of antigenic relationship and acquisition of natural immunity without morbidity, have already been observed with several viruses, including filoviruses and flaviviruses (Figure 2) [9-11].

Figure 2. Examples of chiropteran geographic distributions: Fruit bats in the World (left), and a common insectivorous bat (Hipposideros Caffer) in Africa (right).
The long genome of Coronaviruses mainly encodes four major structural proteins: spike (S); envelope (E); membrane (M); nucleocapsid (N). The spike-shaped transmembrane glycoprotein (S) on the surface of the virus plays an essential role in virus attachment, fusion, entry, and transmission. It comprises two functional subunits: S1 subunit responsible for binding between the virus and the receptor; and S2 subunit (C-terminal stem) that allows fusion of viral and cellular membranes. The S1 subunit is divided into an N-terminal domain (NTD) and a receptor binding domain (RBD) of the C-terminal region responsible for binding the virus to the host cell (ACE2) receptor binding domain [12,13]. The nucleocapsid (N protein) is involved in the packaging of RNA during the externalization of viral particles from the infected cell and is an internal protein of the virus [14]. The N protein appears more conserved across Beta coronavirus species than the S protein, while the RBD appears more conserved within the S1 unit. From the point of view of protection (i.e., neutralizing antibodies), there is a strong correlation between the levels of RBD antibodies and the neutralizing antibodies to SARS-CoV-2 in humans [15]. Also, SARS-CoV-2 genome is closely related to SARS-CoV-1 (79.6%) genomic sequence identity, several antibodies covering all structural proteins of SARS-CoV-1 (spike, membrane, nucleocapsid, envelope) have been identified and extensively studied showing cross-reactivity with SARS-CoV-2, as well as partial crossneutralization of spike antibodies [16]. Moreover, sera from SARS-CoV-1 convalescent or S1 CoV-specific animal antibodies can neutralize SARSCoV- 1 infection by reducing S protein-mediated SARS-CoV-1 entry [17]. Finally, SARS-CoV-1 and MERS-CoV show that many fragments (S1-NTD, RBD, S2) of S protein are targets for neutralizing antibody production [18].
Antibody detection
The INNOBIOCHIPS ELISA serological test used detects the IgG antibodies targeting the N protein, the S1 protein, the RBD domain of the S1 protein, the NTD domain of the S1 protein, and the S2 protein [19] from SARS-CoV-2 virus (Wuhan strain). The test values are obtained by optical density reading using a laser reader. The specificity of the test was evaluated using 25 samples positive for low pathogenic human coronaviruses (229E, OC43, NL63, HKU1) [20]. All 25 samples tested negative for all SARSCoV- 2 proteins used in the ELISA test.
Data analysis
After calculating the statistical moments and the distribution of the samples values for each antigen, several statistical tests and calculations were performed including: for each antigen, comparison of the means (Student's T-test) and the variances (F-test) between the samples group and the "control" group; difference between the two groups (samples and controls) considering all the five antigens together was tested using Hotelling test; the control group was taken as a whole, without excluding the few suspected false-negative samples. Calculation of the number and percentage of samples considered positive for each antigen (with confidence interval), according to the cut off value; calculation of the number of positive samples for two or more antigens.
All data are available in the main text or the supplementary materials (2021-Results PRECOV Africa.xlsx).
Control sera collection
The controls sera were obtained by INNOBIOCHIPS Company from 189 samples from blood donors collected in Northern France before the pandemic, randomly selected (Etablissement Français du Sang) and tested negative for SARS-CoV-2 by PCR. These sera collection was used by the manufacturer to define the thresholds of positivity as compared to the sera collected from patients infected by SARS-CoV-2 (PCR test positive). We used this control group to establish thresholds for the absence of SARSCoV- 2-like antibodies with respect to their geographic origin while such blood donors from France are supposed to have not been in direct or indirect contact with bats. To eliminate the risk of false negatives (samples may came from donors of African or South and South-East Asian origin), for each antigen the distribution of control values was modeled. For each antigen, the PRECOV threshold value corresponds to a probability equal to 0.0002. All control samples with a value for an antigen above the threshold will be considered as false negatives for this antigen.
Sample sera collection
We tested a total of 1655 samples from DRC (Democratic Republic of the Congo), Congo, Cameroon and Senegal. DRC samples originated from the Monkole Hospital Center bio bank (190 samples, collected in 2019 from healthy subjects from the hospital staff, from volunteers, and from young sickle-cell disease patients who are part of a study cohort), and from the ALTADEVA/Monkole bio bank (384 samples, collected in 2014 and 2015 as part of a study of Plasmodium falciparum chemo resistance in the cityprovince of Kinshasa, in the central province of Kongo, and south western DRC).
The 383 tested samples from Cameroon were selected among samples received from various laboratories between June 2018 and June 2019 at the Chantal BIYA International Research Centre for HIV Prevention and Management (CIRCB), as part of continual health monitoring among PLHIV. 51% of the 383 samples selected were among samples received from some Central and General Hospitals within the country. The remaining 49% were selected among samples received from peripheral healthcare facilities.
The samples from the Republic of Congo (536 tested samples) were collected by The Fondation Congolaise pour la Recherche Médicale in Southern district of Brazzaville, Madibou and in the Northern part of the country (Sangha) in the district of Bomassa, in 2016 and 2019, respectively.
The samples from Senegal (162 samples) were randomly selected from serums collected in 2018 and 2019 and conserved in the bio bank of the Institute de Recherche en Sciences de la Santé, Epidémiologie et formation (IRESSEF, Health Sciences Research Institute of Epidemiology and Training).
All samples were aliquoted and kept frozen as appropriate and each sample had companion data including date of collection, age, sex, and province of origin.
Ethical approvals
All samples used in this study were collected before 2020 in the laboratories of the partner institutes from volunteer donors for diagnostic purposes. Blood samples were collected after informed consent for the use and reuse from each patient or from his or her parent/guardian in the case of minors. All experiments were performed in accordance with relevant named guidelines and regulations. All documents and samples were anonymized. For samples from DRC, ethical approval was obtained from the Centre de Formation et d'Appui Sanitaire/Centre Hospitalier Monkole ethic committee. For samples from Republic of Congo, ethical approval by the Fondation Congolaise Pour la Recherche Médicale (Congolese Foundation for Medical Research) Institutional Review Board (IRB) and ethic committee. For samples from Cameroon, full approval was obtained from the IRB of the Centre International de Référence Chantal Biya (Chantal Biya International Reference Center). Full approval was obtained from the IRESSEF IRB for the Senegalese samples.
Our results are in favor of the hypothesis that some populations in Africa and potentially from other part of the World might be less susceptible to the SARS-CoV-2 infection due to a pre-existing immunity triggered by other not yet identified Beta coronavirus of animal origin. Indeed, several Beta coronaviruses closed to the Sarbecovirus cluster have been found in horseshoe bats, as well as specific antibody response to these viruses in African fruit bats [21,22]. A new bat sarbecoviruses isolated from Microchiroptera (i.e., insectivorous bats) in Laos seems to have the same potential for infecting humans as early strains of SARS-CoV-2 and for being, so far, the closest strain phylogenetically [23]. This leads one to think by analogy that the African populations could also be exposed to an African Microchiroptera Sarbecovirus, which is yet to be discovered, knowing that these bats are also very common on this continent [8]. Moreover, coronaviruses were detected from several bat species in close contact with human in Rwanda (Africa) including know and novel Beta coronaviruses genetically close to the SARS-CoV-2 [24].
Higher serological cross-reactivity to SARS-CoV-2 in sub-Saharan African regions than else where has already been reported and attributed to higher exposure to human coronaviruses (HCoVs). Although it has been showed that cross-reactive T cells against SARS-CoV2 can be induced by common cold coronavirus, SARS-CoV-1 virus or eventually other animal Beta coronaviruses [25-27], this is the first time that specific antibodies against SARS-CoV2 proteins in sera collected before the SARS-CoV-2 emerged in China have been reported in Africa. The novelty of our finding is that a stronger cross-reactivity in some African regions may come from exposure, not to HCoVs as already published [28], but to other animal coronaviruses circulating in these areas. Indeed, the specificity of the ELISA used in this work excludes reactions against congenital human coronaviruses (e.g., HCoV-NL63, HCoV-229E) and confirms that the identified antibodies react precisely against the SARS-Cov-2 proteins.
We observed a stronger cross-reactivity to S1, S2 and RBD (considered specific to SARS-CoV-2) than for the N protein, considered common to Beta coronaviruses. Indeed, while the N protein is the most conserved, the S1 subunit is the least conserved and cross-reactivity cannot be explained by exposure to the known human CoVs. Therefore, it can be assumed that this cross-reactivity must have been induced by a virus close (i.e., sharing spike epitopes) to SARS-CoV-2, rather than by other known human Beta coronaviruses. Moreover, such cross-reactivity with S1 does not quantitatively reproduce the S1-RBD or S1-NTD data. These consistently observed discrepancies can be explained by a variation of antibodies affinities to these epitopes due to a change of, either or both, the structure of the spike or their amino-acid sequence. Eventually a more consistent response with S1 could be the fact of a non-tested here CTD protein [29]. Altogether, such discrepancy of antibody response is in favour of an S1 belonging to a SARS-CoV-2-like virus, while all sequence of the antigens included in the commercial ELISA (INNOBIOCHIPS) were entirely based on the reference SARS-CoV-2 strain. It is also important to note that S and N protein sequences are equally divergent among coronaviruses while the S2 subunit is better conserved than the N protein. Moreover, with respect to new bat sarbeco viruses isolated in Laos the authors showed that the spike of SARS-CoV-2 is a mosaic of sequences extremely close to the one of this new bat virus isolated [23]. Ultimately the RBDs of these viruses slightly differ from that of SARS-CoV-2 and bind as efficiently to the hACE2 protein as the SARS-CoV-2 Wuhan strain isolated in early human cases. Moreover, it is established that the RBD as a hACE2-dependent mediator in human cells is inhibited by antibodies neutralizing SARS-CoV-2 [30].
We are aware of the limitations of this somewhat pioneering study. The next step is to conduct sensitive investigations among populations potentially exposed to wild animals and to perform back-to-back tests with the present ELISA test and with essential SARS-CoV-2 neutralization assays to confirm the surprisingly high S1 cross-reactivity and the potential of the detected pre-pandemic antibodies to protect against SARS-CoV-2. Furthermore, if it is true that background reactivity in SARS-CoV-2 serological tests is higher, especially in central African populations, this may be due not only to widespread circulation/exposure of/to Betacoronavirus of chiropterans or other animals, but to a potential increased cross-reactivity induced by other microorganisms (e.g., malaria, tuberculosis, etc.) as previously observed [31-33].
The controls samples from France blood donors tested for SARSCoV- 2 reacting antibodies are shown in Table 1. Among 189 control samples, we detected 18 samples with antibodies reacting against at least 1 antigen (9.5%), and none sample with antibodies reacting against at least 2 antigens (0%).
| Antigen (189 samples) | N | S1 | S2 | S1-RBD | S1-NTD |
|---|---|---|---|---|---|
| Min | 0 | 0 | 0 | 0 | 0 |
| Max | 17.95 | 1.40 | 13.56 | 1.10 | 5.24 |
| Mean | 0.695 | 0.106 | 1.125 | 0.105 | 0.353 |
| 1st Quartile | 0.07 | 0.04 | 0.08 | 0.02 | 0.03 |
| Median | 0.117 | 0.067 | 0.254 | 0.067 | 0.114 |
| 3rd Quartile | 0.31 | 0.10 | 1.13 | 0.12 | 0.28 |
| Standard Deviation | 2.22 | 0.16 | 2.11 | 0.158 | 0.796 |
| PRECOV Threshold | 9 | 0.6 | 10 | 0.6 | 4.5 |
| Value above threshold* | 1.6%(3) | 2.1%(4) | 1.5%(3) | 2.6%(5) | 1.6%(3) |
Note: ⁎=Percentage (sample size above threshold); RBD: Receptor Binding Domain; NTD: N-Terminal Domain
Antibodies against the five tested SARS-CoV-2 antigens were detected in the pre-COVID samples, with differential optical density mean value for African samples significantly higher than for the control samples. The S1 antigen shows the highest percentage of positives: 19.64% for African samples versus 2.11% for the control samples. The S2 and RBD antigens also show significantly higher rates. We find also a high significant difference with control samples (p-value<10-6) when all antigens were considered together. Among the 1655 tested samples, 630 samples reacted at least against 1 antigen above the threshold (38.1%, vs. 9.5% for the controls), while 205 samples reacted at least against 2 antigens above the threshold (12.4%, vs. 0% for the controls) (Table 2).
| Results | N | S1 | S2 | S1-RBD | S1-NTD |
|---|---|---|---|---|---|
| Minimum | 0 | 0 | 0 | 0 | 0 |
| Maximum | 59.09 | 50.28 | 95.40 | 45.34 | 70.37 |
| Mean | 2.63 | 0.98 | 4.39 | 0.44 | 1.19 |
| Standard Deviation (SD) | 6.86 | 3.79 | 10.65 | 2.11 | 4.40 |
| 1st quartile | 0.13 | 0.08 | 0.12 | 0.005 | 0.03 |
| Median | 0.53 | 0.17 | 0.59 | 0.08 | 0.13 |
| 3rd Quartile | 1.63 | 0.47 | 2.86 | 0.23 | 0.44 |
| Total$ | 6.77 [5.6-8.0] (112) | 20.18 [18.2-22.1] (334) | 10.75 [9.3-12.4] (178) | 11.05 [9.5-12.6] (183) | 5.50 [4.4-6.6] (91) |
| Student T-test | p#=6*10-5 | p=7*10-4 | p=1.4*10-5 | p=0.0159 | p=0.0046 |
Note: $=Percentage of samples above threshold, [95% CI], (number); #=p-value; RBD: Receptor Binding Domain; NTD: N-Terminal Domain
The results by country Table 3 show higher values for Cameroon, especially for S1, S2, and S1-RBD antigens. The other antigens do not show significantly different values between countries. All values are significantly higher than control values.
| Country | N | S1 | S2 | S1-RBD | S1-NTD |
|---|---|---|---|---|---|
| Cameroon (383)* | 5.7% [3.4-8.0] |
32.4% [27.7-37.1] |
18.0% [14.2-21.8] |
22.2% [17.9-26.1] |
6.0% [3.6-8.4] |
| Congo (ROC) (536)* | 7.6% [5.4-9.8] |
13.6% [10.7-16.5] |
10.8% [8.2-13.4] |
8.8% [6.4-11.2] |
5.2% [3.3-7.1] |
| Congo (DRC) (574)* | 7.3% [5.2-9.4] |
19.2% [16.0-22.4] |
7.8% [5.6-10.0] |
5.7% [3.8-7.6] |
5.6% [3.7-7.5] |
| Senegal (162)* | 4.3 % [1.2-7.4] |
16.6% [10.9-22.3] |
3.7% [0.8-66] |
11.1% [6.3-15.9] |
4.9% [1.6-8.2] |
Note: ⁎=(sample size); RBD: Receptor Binding Domain; NTD: N-Terminal Domain
Sera samples from blood donors of France also collected before the pandemic were used as control. The results showed a statistically highly significant difference for antibodies prevalence between the samples collected in Africa and the control samples. Our results suggest that in the tested African sub-regions the populations have been potentially preexposed before the COVID-19 pandemic to the antigens of a SARS-CoV- 2-like virus.
Virology: Current Research received 187 citations as per Google Scholar report