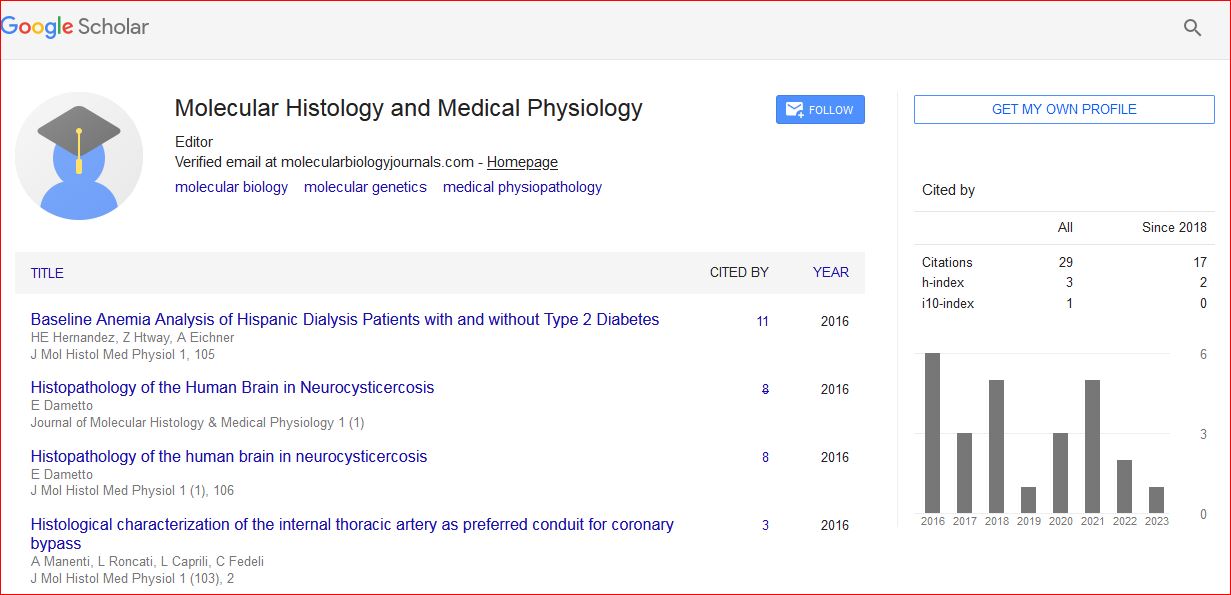
Mini Review - (2022) Volume 7, Issue 5
Received: 05-Sep-2022, Manuscript No. jmhmp-23-86953;
Editor assigned: 07-Sep-2022, Pre QC No. P-86953;
Reviewed: 19-Sep-2022, QC No. Q-86953;
Revised: 26-Sep-2022, Manuscript No. R-86953;
Published:
01-Oct-2022
, DOI: 10.37421/2684-494X.2022.7.50
Citation: Arpi, Darshini. “Structure of Nucleus in Smooth Muscle Cell.” J Mol Hist Med Phys 7 (2022): 50.
Copyright: © 2022 Arpi D. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Smooth muscle is a kind of tissue that is present in the walls of hollow organs like the stomach, uterus, and intestines. Additionally, smooth muscle is present in the walls of arteries and veins in the cardiovascular system. The urinary, pulmonary, and reproductive tracts all contain this kind of involuntary non-striated muscle. Additionally, smooth muscle can be found in the eyes, where it alters the shape and size of the lens and iris. Additionally, the skin has smooth muscle, which enables hair to stand up in response to cold or fear.
Muscle fibers • Uterus • Cytoplasm
Fructose 1,6-bisphosphatase catalyses the hydrolysis of fructose 1,6-bisphosphate to fructose 6-phosphate and inorganic phosphate in the presence of divalent metal ions (Mg2+, Mn2+, Co2+, and Zn2+). The enzyme was first discovered by Gomori. Over the following 60 years, almost 2000 papers investigating the kinetics and tissue distribution of FBPase were released. However, only a small number of these investigations focus on the subcellular localization of the enzyme. Saez et al. first identified FBPase in the perinuclear region of hepatic and renal cells [1], first identified FBPase's nuclear localization in these cells. According to a recent investigation of the subcellular distribution of the enzyme in mammalian muscle tissue, FBPase is present on both sides of the Z-line in skeletal muscle and interacts extensively with a-actinin. It is also identified in the nuclei of cardiac muscle cells. In this study, we demonstrate that FBPase is distributed subcellularly in smooth muscle cells similarly to cardiomyocytes and that the enzyme is only found in the heterochromatin region of the cells' nucleus. In vertebrate tissues, two FBPase isozymes have been found [2]. A liver enzyme that controls gluconeogenesis is called FBPase. The muscle isozyme participates in both the regulation of glycolysis and the synthesis of glycogen from lactate. The presence of the enzyme in the cell nucleus is not necessary for any of these reactions. In order to find a molecular basis for FBPase's nuclear localization, we constructed the structures of every known muscle FBPase sequence. Using in silico analysis of the tertiary structures, we found a number of functional sites suggestive of a wide range of proteins amenable to nuclear transport. The physiological relevance of this observation is investigated. Fluka supplied the paraffin wax, paraformaldehyde, glutaraldehyde, Coomassie Brilliant Blue R-250, and antibody diluent; DAKO supplied the DAB chromogen; and ICN supplied the anti-rabbit IgG goldconjugated (10 nm) antibody. The ExtrAvidin Horseradish Peroxidase, Normal Sera, Nitrocellulose Membranes, Biotin Conjugated Mouse Monoclonal Anti- Rabbit Immunoglobulins, and other reagents were given by Sigma. All of the reagents were of the maximum purity that was commercially feasible [3]. By injecting the electrophoretically pure enzyme and Freund's complete adjuvant into the rabbit's skin, muscle-specific antiserum FBPase was produced. The experiment was conducted in compliance with the guidelines set forth by The Scientific Research Ethical Committee. The immunoserum was partially purified using acetone powder. The reactivity of the anti-FBPase serum was evaluated using the twofold diffusion technique. After immunodiffusion, the antiserum and the purified enzyme both responded strongly [4].
The smooth muscle cell is 20–200 m length and 3–10 m thick. All of the cytoplasm is uniformly eosinophilic, and myofilaments make up the majority of it. Located in the middle, the nucleus resembles a cigar as it contracts. Caveolae, which are similar in function to the T-tubules of the skeletal muscle, are tiny pouch-like invaginations of the cell membrane into the cytoplasm. A basal lamina serves as the smooth muscle cells' anchor to the surrounding connective tissue. Branching bundles of smooth muscle fibres form a group. In contrast to skeletal muscle fibres, these bundles form a complicated structure rather than running rigidly parallel and in an organised fashion. The cells can therefore contract much more forcefully than striated muscle. Between dense bodies in the cytoplasm and attachment plaques at the cell membrane, the actin filaments are stretched. Between the actin filaments are the myosin filaments. Furthermore, the cell structure is supported by intermediate filaments like desmin and vimentin [5].
One can distinguish between single-unit and multi-unit types functionally. The single-unit kind of smooth muscle cells contract uniformly and are electrically coupled by gap junctions. The blood arteries and internal organs' walls include these cells (visceral smooth musculature). Because the multi-unit smooth cells are separate from one another, each one must be innervated separately to enable more precise muscle control. They can be found in the iris and hair erector muscles, among other places.
None.
None.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at