Opinion - (2022) Volume 13, Issue 7
Received: 02-Jul-2022, Manuscript No. jar-22-73705;
Editor assigned: 04-Jul-2022, Pre QC No. P-73705;
Reviewed: 16-Jul-2022, QC No. Q-73705;
Revised: 21-Jul-2022, Manuscript No. R-73705;
Published:
28-Jul-2022
, DOI: 10.37421/2155-6113.2022.13.897
Citation: Kent, Stephen. “HIV Concealment by Have Limitation Factors and Viral Insusceptible Avoidance.” J AIDS Clin Res 13 (2022): 897.
Copyright: © 2022 Kent S. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Antiviral restriction factors are an integral part of the host innate immune system that protects cells from viral pathogens, such as human immunodeficiency virus (HIV). Studies of the interactions between restriction factors and HIV have greatly advanced our understanding of both the viral life cycle and basic cell biology, as well as provided new opportunities for therapeutic intervention of viral infection. Here we review the recent developments towards establishing the structural and biochemical bases of HIV inhibition by, and viral countermeasures of, the restriction factors TRIM5, MxB, APOBEC3, SAMHD1, and BST2/tetherin.
The host innate immune system has a diverse array of antiviral responses, including intrinsic antiviral proteins termed restriction factors that inhibit distinct stages of the viral life cycle. The last decade of HIV research has seen the discovery of many anti-HIV restriction factors that blocks the infection at steps including capsid uncoating (TRIM5), reverse transcription (APOBEC3 and SAMHD1), nuclear import and integration (MxB), translation (Schlafen 11), and budding (BST2/Tetherin). Some of these restriction factors also trigger broad innate immune signaling for further viral suppression. The virus, in turn, has developed an arsenal of methods to evade the host defense. HIV counteracts or avoids some of these restriction factors either by using antagonistic accessory proteins (Vif, Vpu, Vpx/Vpr, Nef, etc.) or by mutating protein interfaces. Studying the ongoing arms race between HIV and the host greatly advances our understanding of the viral life cycle and the human immune system, while simultaneously providing new avenues for therapeutic intervention [1,2].
The tripartite motif 5 (TRIM5) proteins, including TRIM5α and TRIMCyp, directly recognize the retroviral capsid in a species-specific manner. TRIM5 proteins elicit premature disassembly of the capsid and activate cellular innate immune signalling pathways. The conserved N-terminal tripartite motif of TRIM5 proteins consists of a RING domain (E3 ligase activity), a B-box domain (higher-order oligomerization), and a coiled-coil domain (dimerization). The C-terminal PRY/SPRY (TRIM5α) or CypA (TRIMCyp) domain confers capsid binding and specificity. Functioning as a viral capsid pattern sensor, TRIM5α binds only to the assembled hexameric capsid lattice. Interestingly, although the global capsid pattern is the binding determinant, TRIM5α has an ability to interact with retroviral capsids of diverse shapes and curvatures [3,4].
Crystal structures of the rhesus macaque TRIM5α PRY/SPRY domain provided the first insight into the potential mode of interaction between TRIM5α and the viral capsid. The structures show a core of two antiparallel β sheets with one side of the molecule displaying flexible variable loops that are critical for capsid binding. Rhesus TRIM5α, but not human TRIM5α, recognizes and restricts HIV-1, although a single mutation (R332P) in the variable loop 1 of the human PRY/SPRY domain confers HIV-1 capsid binding and restriction. Modelling of the TRIM5α PRY/SPRY domain onto the capsid lattice suggests that the flexibility provided by the variable loops may enable TRIM5α to recognize retroviral capsids with a variety of curvatures [5].
The TRIM5 B-box and coiled-coil domains mediate higher-order selfassociation that enables avid binding to retroviral capsid cores, as monomeric TRIM5 domains bind capsid weakly. A TRIM5-TRIM21 chimera (TRIM5- 21R) has been observed by electron microscopy to form a two-dimensional hexagonal lattice on the surface of a preassembled HIV-1 capsid lattice. The crystal structures of the coiled-coil region of TRIM25 and the B-box-coiledcoil region of TRIM5α provided insights into TRIM5 dimerization and lattice assembly. The coiled-coil region of TRIM proteins adopts an antiparallel conformation, with a downstream α-helix folding back in a hairpin-like manner to bring the capsid-binding modules near the midpoint of the coiled-coil. The length of the coiled-coil (17 nm) matches the dimension of the observed TRIM5- 21R hexagonal lattice. The TRIM5α B-box domains sit at opposite ends of the coiled-coil. This configuration suggests that within the TRIM5 lattice the B-box domains locate at three-fold symmetry axes, while each PRY/SPRY or CypA pair reside at the center of a hexagonal edge for viral capsid interaction.
Structural studies of more complete TRIM5 constructs containing the coiled-coil and additional domains (RING, PRY/SPRY or CypA) are needed to further understand TRIM5 architecture. More importantly, the most intriguing and challenging questions remain: what is the structural basis of capsid lattice recognition and disruption by TRIM5α, and how does this recognition lead to downstream immune responses? Answering these questions will likely require innovative approaches to reconstitute the lattice interactions in a form amenable for biochemical and structural biology studies [6].
Human MxB is an interferon-induced restriction factor, which was recently discovered to target HIV-1. Evidence suggests it acts between reverse transcription and integration, and likely interacts with the viral capsid. MxB is highly homologous to the well-studied MxA (63% identity) that inhibits influenzalike viruses. Both MxA and MxB are dynamin-like GTPases that contain three domains: GTPase, bundle signaling element (BSE), and stalk. The recent crystal structure of MxB shows an MxA-like extended anti-parallel dimer. The relative orientations of the structurally homologous domains are different in the two proteins, although the functional significance of this difference is currently unclear.
The antiviral mechanism of MxB is distinct from that of MxA. MxB, but not MxA, contains a nuclear localization signal (NLS), which is critical for HIV- 1 restriction. The anti-HIV-1 function of MxB is independent of the transfer of information between the GTPase and the stalk domains through hinge regions surrounding the BSE domain. This is consistent with data showing that MxB antiviral function is independent of GTPase activity. Both BSE hinge communication and GTPase activity are critical for MxA antiviral functions. In addition, higher-order oligomerization is required for the antiviral activity of MxA but not MxB. Strikingly, the viral interaction modules of MxA and MxB appear to lie on the opposite faces of the dimer structures.
MxB binds to HIV-1 capsid assemblies, but not to capsid protein (CA) hexamers, indicating that MxB may function as another viral capsid pattern sensor. The MxB regions critical for antiviral function, such as the N-terminus and the dimerization interface, are also important for capsid binding. Intriguingly, MxB also interacts with CA mutants that evade MxB restriction in vivo, indicating capsid binding by MxB may be necessary but not sufficient for HIV-1 restriction. The current data suggest that MxB may restrict HIV-1 infection by interfering with viral nuclear import. However, much remains to be uncovered to explain the detailed mechanism of this newly discovered restriction factor.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
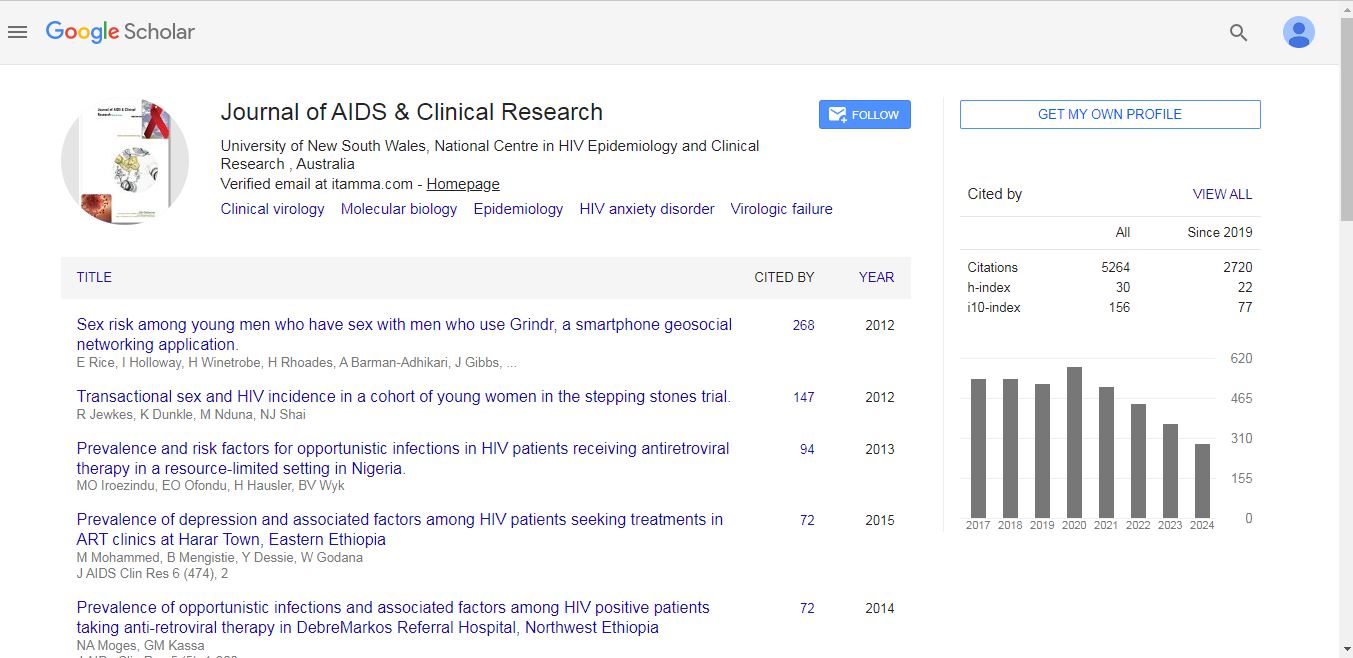
Journal of AIDS & Clinical Research received 5264 citations as per Google Scholar report