Editorial - (2021) Volume 5, Issue 4
Received: 08-Jul-2021
Published:
21-Jul-2021
, DOI: 10.37421/jmbp.2021.5.126
Citation: Xi-he Zhang. "Pathotypes of Enteropathogenic E. coli (EPEC)." J Microbiol Pathol 5 (2021): 126.
Copyright: © 2021 Xi-he Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited
EPEC was the primary pathotype of E. coli to be described. Large outbreaks of infant diarrhoea within the uk led Bray, in 1945, to explain a group of serologically distinct E. coli strains that were isolated from children with diarrhoea but not from healthy children. Although large outbreaks of infant diarrhoea thanks to EPEC have largely disappeared from industrialized countries, EPEC remains a crucial explanation for potentially fatal infant diarrhoea in developing countries 2. for many years, the mechanisms by which EPEC caused diarrhoea were unknown and this pathotype could only be identified on the idea of O:H serotyping. However, since 1979, numerous advances in our understanding of the pathogenesis of EPEC diarrhoea are made, such EPEC is now among the simplest understood of all the pathogenic E. coli.
A characteristic intestinal histopathology is related to EPEC infections; referred to as 'attaching and effacing' (A/E), the bacteria intimately attach to intestinal epithelial cells and cause striking cytoskeletal changes, including the buildup of polymerized actin directly beneath the adherent bacteria. The microvilli of the intestine are effaced and pedestal-like structures on which the bacteria perch frequently get up from the somatic cell, the power to induce this A/E histopathology is encoded by genes on a 35-kb pathogenicity island (PAI; see below) called the locus of enterocyte effacement (LEE). Homologues of LEE also are found in other human and animal pathogens that produce the A/E histopathology, including EHEC, rabbit EPEC (REPEC) and Citrobacter rodentium, which induces colonic hyperplasia in mice. The LEE encodes a 94-kDa outer-membrane protein called intimin, which mediates the intimate attachment of EPEC to epithelial cells. Intimin not only functions as a ligand for somatic cell adhesion, but also stimulates mucosal TH1 IMMUNE RESPONSES and intestinal crypt hyperplasia. Most of the 41 open reading frames of the core LEE PAI encode a kind III secretion system and therefore the associated chaperones and effector proteins. one among these effector proteins, referred to as Tir (translocated intimin receptor), is inserted into the host-cell membrane, where it functions as a receptor for the intimin outermembrane protein. this is often a desirable example of a pathogen that gives its own receptor for binding to eukaryotic cells, although additional eukaryotic proteins have also been reported to act as receptors for intimin. A recent study showed that EPEC can disrupt cell polarity, causing basolateral membrane proteins, especially β1-integrins, to migrate to the apical cell surface where they will bind to intimin. additionally to β1-integrin, Tir has also been shown to bind to NUCLEOLIN. additionally to its role as a receptor for intimin, Tir has important signalling functions in epithelial cells. The portion of Tir that's exposed to the cytosol nucleates cytoskeletal proteins, initially binding on to the adaptor protein Nck, which recruits the amino terminus of Wiskott–Aldrich syndrome protein (N-WASP) and therefore the actin-related protein 2/3 (Arp2/3) complex; recruitment of Arp2/3 leads to actin filament nucleation and initiation of the characteristic pedestal complex. Interestingly, the Tir protein of EHEC O157:H7 isn't functionally just like the Tir protein of EPEC O127:H6 because pedestals are formed independently of Nck, which indicates that additional bacterial factors are translocated to trigger actin signalling. Other cytoskeletal proteins, like vinculin, cortactin, talin and α-actinin, also are recruited to the pedestal complex. Formation of the pedestal may be a dynamic process whereby the force of actin polymerization can propel the pedestal across the surface of ptK2 epithelial cells. Tir also features a GAP (GTPase-activating protein) motif that has been implicated within the ability of Tir to downregulate filopodia formation. Another secreted effector protein is EspF, which causes apoptosis and induces redistribution of the tight-junctionassociated protein occludin, which results in loss of trans-epithelial electric resistance. As noted above, the Map protein affects mitochondrial function and filopodia formation, and extra effectors — for instance, EspG and EspH.
EPEC virulence factors that are encoded outside the LEE have also been described. One very large protein of ∼385 kDa called lymphostatin (LifA) inhibits lymphocyte activation 33. This protein is additionally present in strains of EHEC, where it's referred to as Efa1, and an adhesive property has been attributed thereto. Typical EPEC strains possess a plasmid of 70–100 kb called the EAF (EPEC adherence factor) plasmid. This plasmid encodes a kind IV pilus called the bundle-forming pilus (BFP), which mediates interbacterial adherence and possibly adherence to epithelial cells. It also contains the per locus (plasmid-encoded regulator), the products of which regulate the bfp operon and most of the genes within the LEE by the LEEencoded regulator (Ler). So-called atypical EPEC contain the LEE but don't contain the EAF plasmid. In industrialized countries, atypical EPEC are more frequently isolated from diarrhoeal cases than are typical EPEC that contain the EAF plasmid, although typical EPEC dominate in developing countries. Atypical EPEC have also caused large outbreaks of diarrhoeal disease involving both children and adults in industrialized countries.
The model of EPEC pathogenesis is considerably more complex than simple binding to epithelial cells by one adhesin and secretion of an enterotoxin that induces diarrhoea. The emerging model, several aspects of which are reviewed elsewhere, indicates that EPEC initially adhere to epithelial cells by an adhesin, the identity of which isn't yet clearly established; potential candidates include BFP, the EspA filament, flagella, LifA/Efa1 and intimin (by host-cell receptors). the sort III secretion system is then activated and various effector proteins — including Tir, EspF, EspG, EspH and Map — are translocated into the host cell. EPEC binds through the interaction of intimin with Tir inserted within the membrane and various cytoskeletal proteins accumulate underneath the attached bacteria. Protein kinase C (PKC), phospholipase Cγ, myosin light-chain kinase and mitogen-activated protein (MAP) kinases are activated, which results in several downstream effects, including increased permeability thanks to loosened tight junctions. Nuclear factor (NF)-κB is activated, resulting in production of IL-8 and an inflammatory response that involves transmigration of polymorphonuclear leukocytes (PMNs) to the lumenal surface and activation of the adenosine receptor. The galanin-1 receptor is upregulated, thereby increasing the response of the epithelial cells to the neuropeptide GALANIN, which is a crucial mediator of intestinal secretion. Some, but not all, typical EPEC strains produce an enterotoxin, EspC, that increases short current in USSING CHAMBERS. Diarrhoea probably results from multiple mechanisms, including active ion secretion, increased intestinal permeability, intestinal inflammation and loss of absorptive area resulting from microvillus effacement.
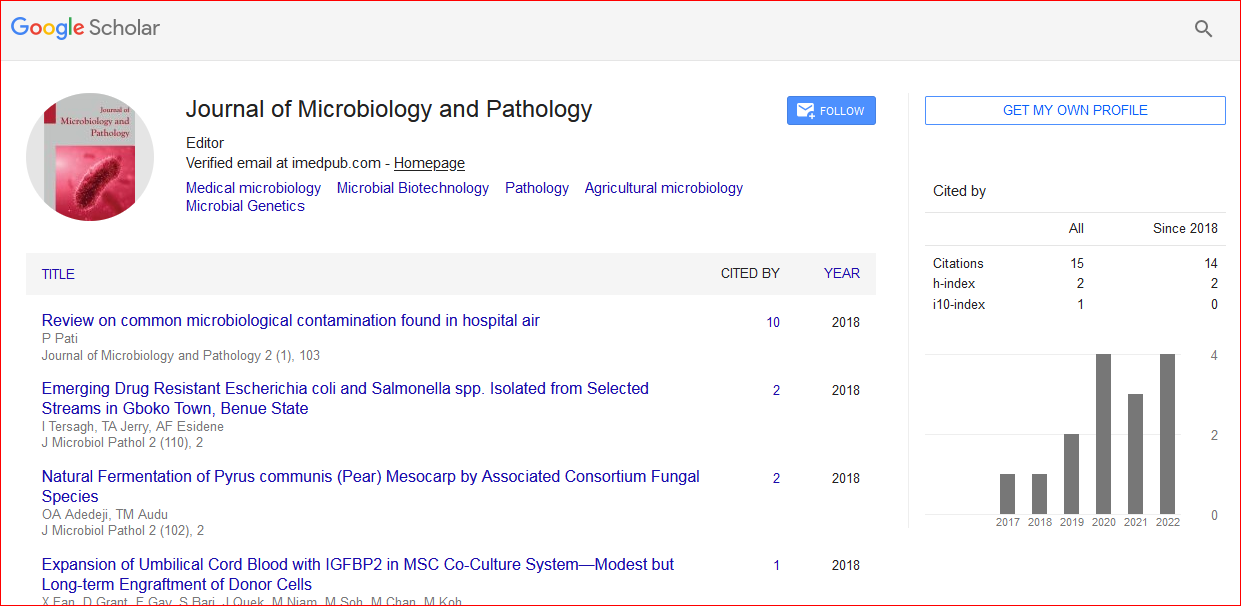
Journal of Microbiology and Pathology received 18 citations as per Google Scholar report