Perspective - (2022) Volume 8, Issue 4
Received: 01-Jul-2022, Manuscript No. jotr-22-76532;
Editor assigned: 04-Jul-2022, Pre QC No. P-76532;
Reviewed: 18-Jul-2022, QC No. Q-76532;
Revised: 24-Jul-2022, Manuscript No. R-76532;
Published:
29-Jul-2022
, DOI: 10.37421/2476-2261.2022.8.206
Citation: Reeves, Bernd. “Analyzing the Costs and Benefits of DNA Repair in Dynamic Situations to Solve the Evolutionary Riddle of Genetic Instability.” J Oncol Transl Res 8 (2022): 206.
Copyright: © 2022 Reeves B. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Hereditary flimsiness assigns different cell aggregates portrayed by an overall expansion in change rate. Such aggregates advance immediately in neoplastic growths and microbial populaces, and assume a basic part in the improvement of disease and anti-toxin obstruction. Hereditary shakiness has been connected with explicit qualities and biochemical pathways. Specifically, microsatellite flimsiness is brought about by lacks in nucleotide confound fix while chromosomal unsteadiness has been connected with surrenders in guideline of chromosomal isolation. As a general rule, it is obvious that hereditary unsteadiness is brought about by long-lasting or inducible lacks in DNA control and fix components and the sub-atomic subtleties are turning out to be progressively clear [1].
The overarching creed is that unsteady cells endure on the grounds that the raised change rate produces versatile variations, and might be alluded to as the transformation for endurance speculation. The essential issue with this model, nonetheless, is that arbitrary transformations are measurably definitely bound to be negative or deadly than they are to be versatile. A few creators have handled this issue by presenting new developmental standards. Advancement by second-request choice, counterselection related determination and mutator catching a ride are various models which express that hereditary flimsiness emerges, not on the grounds that maintenance inadequacy is great for the singular cell, but since the raised change rate builds the populace's general opportunity of endurance. On a basic level consequently, they all make sense of hereditary unsteadiness as an outcome of gathering choice. The idea of gathering choice has been broadly bantered for over a long period and we will here shun further elaborations [2].
Natural development essentially concerns engendering of data as nucleotide groupings or other actual examples like those of DNA methylation. Determination suggests preservation of data, and when we say that a cell has been chosen, it is really shorthand for saying that sub-atomic examples of data have been duplicated from one cell to its relatives. The genome is chosen. In any case, even that is only an estimation. Replication is flawed, and a few sections of the genome are essentially modified. These groupings are by definition not chose. Running against the norm, they are transformed. Stringently talking, thusly not the phone is chosen, just its monitored examples of sub-atomic data. This degree of accuracy might appear to be a piece overexpress. By the by, it is critical to perceive that the idea of cell choice is an estimation. This estimate falls apart with expanding transformation rate, and as we would like to think, it separates with regards to displaying hereditary shakiness. This developmental issue subsequently requests a severe subatomic viewpoint [3].
Hereditary flimsiness is fundamentally brought about by misfortune or hushing of qualities that straightforwardly or in a roundabout way advance tough replication of DNA. These nucleotide arrangements, for the most part alluded to as DNA fix qualities, are the genuinely determinable articles that underlie advancement of hereditary flimsiness. The term DNA fix usually suggests a system pretty much straightforwardly engaged with the acknowledgment and expulsion of DNA harm. On a fundamental level notwithstanding, it is relevant to any cycle that in some way adds to the respectability of the genome. A model of hereditary flimsiness ought to in this manner make sense of why replication loyalty is negative in specific conditions, and as a general rule, we need to consider the developmental upsides and downsides of a DNA fix quality [4, 5].
The developmental benefit of a DNA fix quality ought to be self-evident. Besides the fact that it catalyzes its own union by layout replication, it additionally encodes proteins that guarantee high-constancy replication of its own succession and the whole genome on which it depends. Thusly, it ought to be inclined toward by regular choice. Be that as it may, fix likewise has a huge drawback. Blunder recognition and amendment takes time and consumes energy and the cost of compelling DNA fix is microscopically appeared by its cozy relationship to cell cycle control. With respect to a wide range of data handling, DNA replication consequently includes an inescapable situation among speed and loyalty. In light of these contemplations we, and others, have presumed that DNA fix isn't really a good system. There could be conditions in which DNA fix costs more than the blunders it forestalls. On a fundamental level, a mutagenic climate could discredit the transformative benefit of the maintenance component that endures it, and this clear inconsistency might be figuratively settled by the expression 'Don't stop for fixes in a disaster area.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
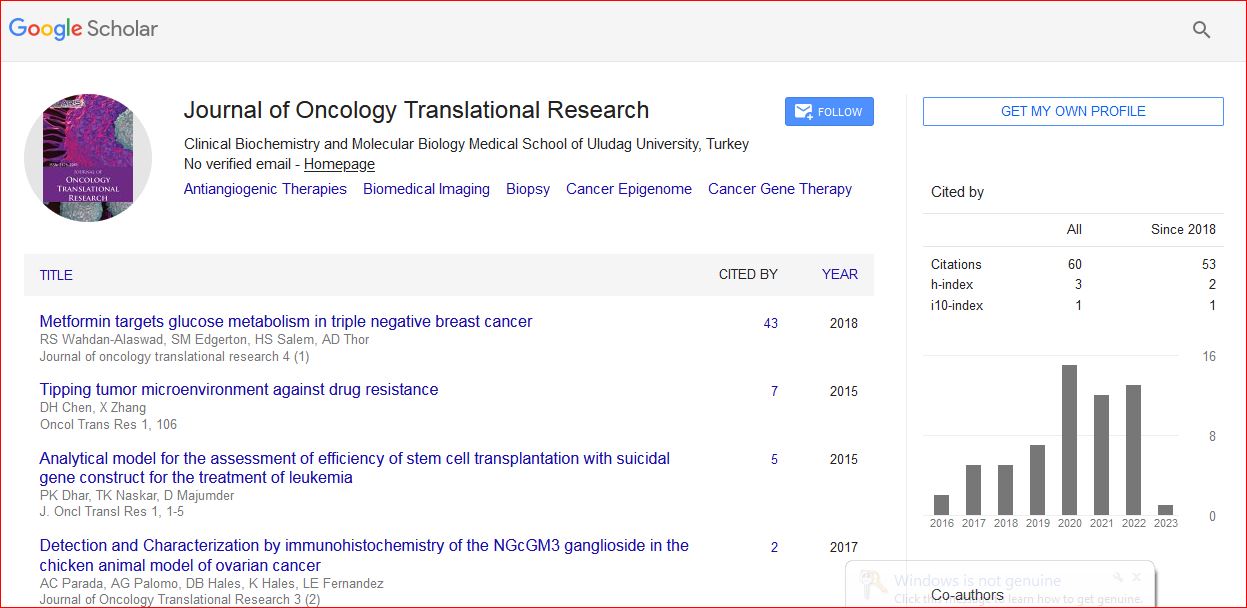
Journal of Oncology Translational Research received 93 citations as per Google Scholar report