Commentary - (2022) Volume 13, Issue 4
Received: 04-Apr-2022, Manuscript No. jch-22-64914;
Editor assigned: 05-Apr-2022, Pre QC No. P-64914;
Reviewed: 18-Apr-2022, QC No. Q-64914;
Revised: 23-Apr-2022, Manuscript No. R-64914;
Published:
30-Apr-2022
, DOI: 10.37421/2157-7099.22.13.627
Citation: Stylli, Nazio. “Regulation of Dynamic Cell Adhesion via Way of Means of Integrin-integrin Crosstalk.” J Cytol Histol 13 (2022): 627
Copyright: © 2022 Stylli N. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Cell adhesion is significant for improvement, the age of organs, cell development and cell dispersion in the body. It connects with grip proteins, which incorporate integrins, immunoglobulin superfamily proteins, starch restricting selectins and cadherins. The integrins are glycoprotein heterodimers shaped by α-and β-chain. They are type 1 film proteins with enormous extracellular spaces, a transmembrane fragment and a cytoplasmic tail. The integrins exist in somewhere around three significant structures. In the resting state, the integrin heads are turned towards the lipid film. Upon initiation, the integrins expand and open the ligand restricting destinations. The integrins are actuated by back to front or outside-in enactment, bringing about intracellular flagging and cell bond. In back to front enactment, an actuating signal starts from a non-integrin receptor, and is sent to integrins. In outside-in actuation, an extracellular ligand ties to an integrin and enacts the integrin. A few cytoplasmic proteins connect with the integrin cytoplasmic tails and are significant in managing integrin movement. When actuated, the integrins may expand their limiting limit by an expansion in partiality of individual integrins through conformational changes, or by grouping of integrins, which is known as expanded devotion [1,2].
Cells can stick to an assortment of cells, and to the extracellular network. To accomplish this, they should communicate a few different integrins with various restricting specificities. These empower them to control bond as per their useful necessities. This is generally clear for portable cells like leukocytes, which experience various cells and extracellular particles, for instance when they leave the veins and enter into tissues. These occasions require the actuation of some integrins, yet additionally the deactivation of others. A few examinations show that integrins impart between one another through intracellular flagging, and through such crosstalk the integrins can collaborate and coordinate cell development and bond in a helpful manner [3]. Besides, different receptors, similar to development factor receptors and infection receptors, may speak with integrins by crosstalk, however in this audit we center around integrin crosstalk. We initially depict how integrins are directed, and the way that they incite intracellular flagging. This data is important to comprehend how integrins impart by crosstalk.
Motile cells should have the option to quickly respond to changes in their current circumstance and stick to their ligands communicated on cells or the extracellular network. This is positively valid for leukocytes and platelets. To get out from the flow into tissues, leukocytes should have the option to tie to endothelial cells, move through the endothelial layer, and connect with extracellular network particles [4]. This implies that they should communicate attachment particles, which can change both the action and explicitness during their movement to the last targets. Protein phosphorylation is the major posttranslational adjustment of proteins, and it empowers proteins to change their movement without the need of new protein union and corruption. The presence of integrin crosstalk might empower the improvement of medications that obstruct the crosstalks. On a basic level, they could be quite certain and bring about less incidental effects. Rather than clearing out all elements of a certain integrin, just a particular cell related capacity would be impacted. By and by, we are just toward the start of a thrilling advancement which during the next few years might change fundamentally [5].
None.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
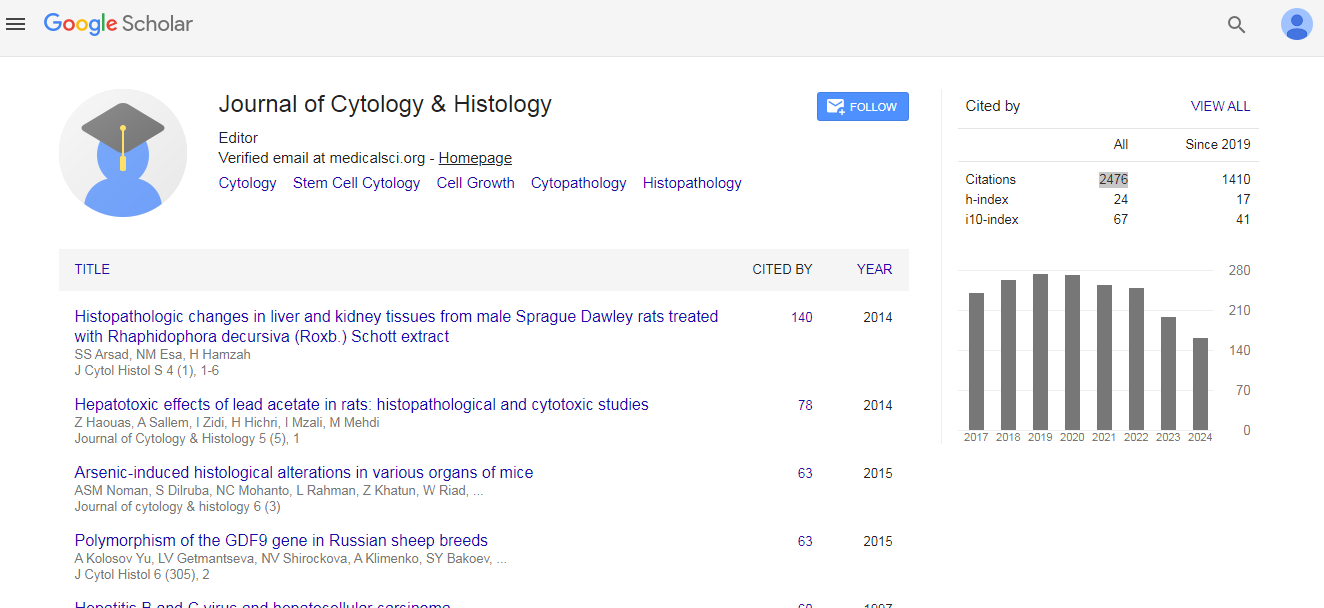
Journal of Cytology & Histology received 2476 citations as per Google Scholar report