Commentary - (2022) Volume 6, Issue 3
Received: 01-May-2022, Manuscript No. JMBP-22-69825;
Editor assigned: 03-May-2022, Pre QC No. P-69825;
Reviewed: 16-May-2022, QC No. Q-69825;
Revised: 21-May-2022, Manuscript No. R-69825;
Published:
28-May-2022
, DOI: 10.37421/2952-8119.2022.6.156
Citation: Chatarjee, Himanshu. “Recognizing and Utilising Plantfriendly Microorganisms.” J Microbiol Pathol 6 (2022): 156.
Copyright: © 2022 Chatarjee H. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Technology advancements and the need for sustainable crop productivity improvements, combined with a century of incremental research, have rekindled interest in the study of advantageous plant-microbe interactions, with a particular focus on how microbiomes affect plant phenotypes. We examine current developments in the study of the plant microbiome and discuss potential implications for boosting crop output [1]. Plant microbiomes' phylogenetic variety is becoming better understood, and it is easier to access their functional diversity. For controlled experiments, there are sizable culture collections available, and more are on the way. On issues relating to microbiome function, genetic resources are being used. In order to promote plant growth and disease resistance, we anticipate that microbial additions of varying complexity will reveal the laws governing these interactions, enabling more sustainable agriculture [2].
Records dating back to 300 BC show that soil microbiome modification to increase crop yield has been done for a long time. Interestingly, the first commercial bioinoculant, nitrogin, was developed in 1896, during the heyday of microbiology and 15 years before the Haber-Bosch method, even though soil microbiomes are currently hailed as a cornerstone of the next green revolution. The Organic Materials Review Institute (OMRI) now classifies 274 goods under the category of "microbial products," either as crop fertilisers or as crop management tools, and 174 products under the category of "microbial inoculants." Since the 1990s, the number of papers relating to microorganisms that promote plant growth (PGP) has increased significantly [3,4].
With the exception of biological nitrogen fixation by rhizobia on legumes and auxin- or ACC-deaminase-mediated phytostimulation, very few of these are linked to mechanistic investigations or modes of action. However, their extensive use has been severely constrained by their narrow host ranges and inconsistent field efficacy. Therefore, it is imperative that we develop a deeper understanding of (a) the mechanisms governing microbial invasion and persistence into standing heterogeneous communities in a variety of habitats, soils, and hosts; and (b) the genetics, present in both partners, that underlies colonisation and the delivery of plant phenotypes by microbes [5].
The emergence of culture-independent microbial ecology, fueled by the development of high-throughput analytic technologies, has made it possible to study the ecological context associated with plants in which microbial inoculants may be used, as well as the mechanisms by which plants regulate the colonisation of beneficial microbes. But to close the current gaps between plant productivity phenotypes and comprehension of the underlying mechanisms, new strategies are required [5]. Because soil microorganisms have a much greater diversity than plant-associated bacteria, it can be deduced that plants are a highly selective microbial niche and that there are broad guidelines for the colonisation of plants by microbes. Read-binning and assembly of bacterial genomes from plant-associated settings can be used as a supplement to shotgun metagenomics to compare the activities of the plantassociated microbiome in the hunt for plant colonisation signals.
Because soil microorganisms have a much greater diversity than plantassociated bacteria, it can be deduced that plants are a highly selective microbial niche and that there are broad guidelines for the colonisation of plants by microbes. To examine the activities of the plant-associated microbiome, read-binning and bacterial genome assembly from habitats associated with plants can be employed as a supplement to shotgun metagenomics to look for signs of plant colonisation.
None.
There is no conflict of interest by author.
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
Google Scholar, Crossref, Indexed at
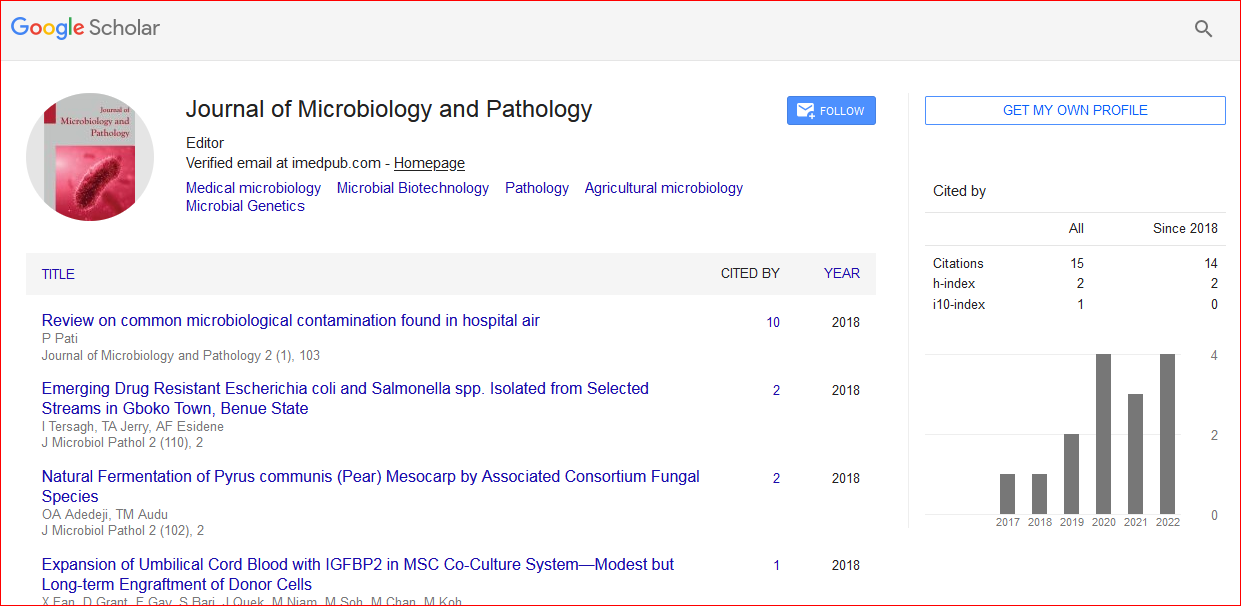
Journal of Microbiology and Pathology received 18 citations as per Google Scholar report