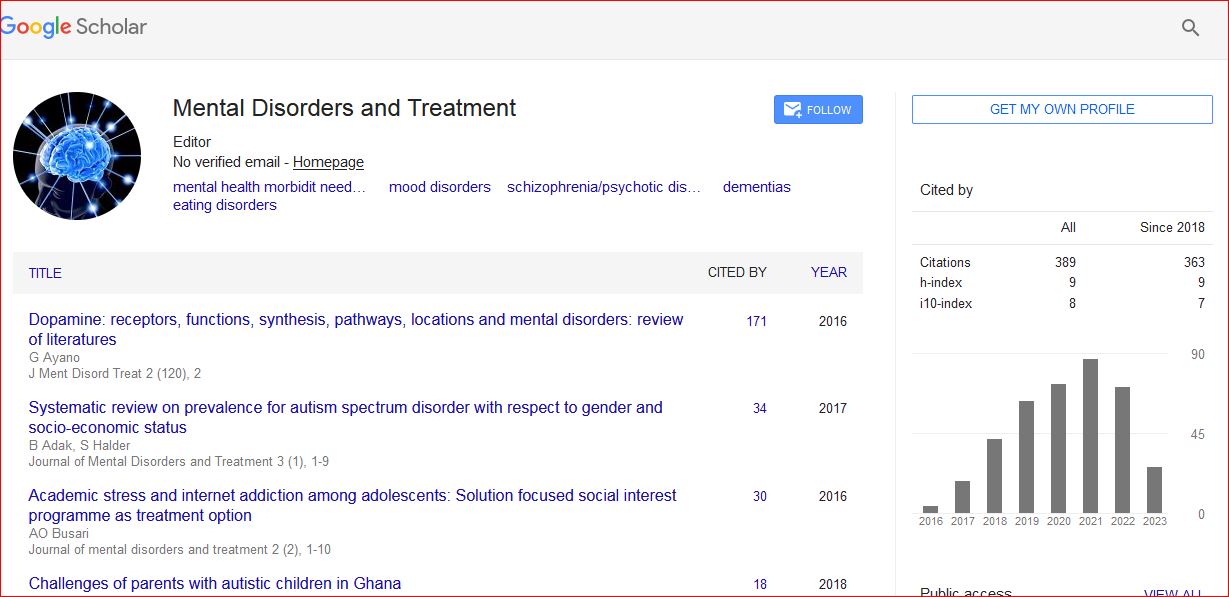
Danielle Meola
Abstract: Brain-derived interleukin-2 (IL-2) has been implicated in diseases processes that arise during CNS development (e.g., autism) to neurodegenerative alterations involving neuroinflammation (e.g., Alzheimer’s disease). Progress has been limited, however, because the vast majority of current knowledge of IL-2’s actions on brain function and behavior is based on the use exogenously administered IL-2 to make inferences about the function of the endogenous cytokine. Thus, to Spot the cell-type(s) and regional circuitry that express brain-derived IL-2, we used B6.Cg-Tg/ IL2-EGFP17Evr (IL2p8-GFP) transgenic mice, which express green fluorescent protein (GFP) in peripheral immune cells known to produce IL-2. We found that the IL2-GFP transgene was localized almost exclusively to NeuN-positive cells, indicating that the IL-2 is produced primarily by neurons. The IL2-GFP transgene was expressed in discrete nuclei throughout the rostral-caudal extent of the brain and brainstem, with the best levels found in the cingulate, dorsal endopiriform nucleus, lateral septum, nucleus of the solitary tract, magnocellular/gigantocellular reticular formation, red nucleus, entorhinal cortex, mammilary bodies, cerebellar fastigial nucleus, and posterior interposed nucleus. Having identified IL-2 gene expression in brain regions related to the regulation of sensorimotor gating (e.g., lateral septum, dorsal endopiriform nucleus, entorhinal cortex, striatum), we compared prepulse inhibition (PPI) of the acoustic startle reaction in congenic mice bred in our lab that have selective loss of the IL-2 gene in the brain versus the peripheral immune system, to test the hypothesis that brain-derived IL-2 plays a role in modulating PPI. We found that congenic mice void of IL-2 gene expression in both the brain and the peripheral immune system, exhibited a modest alteration of PPI. These finding suggest that IL2p8-GFP transgenic mice is also a a useful tool to elucidate further the role of brain-derived IL-2 in normal CNS function and disease.
Keywords: Interleukin-2, Congenic mice, Sensorimotor gating, Prepulse inhibition
Introduction:
Research has implicated both peripheral immune and brain-derived interleukin-2 (IL-2) in neurologic disease processes that arise during CNS development (e.g., autism) to neurodegenerative alterations involving neuroinflammation (e.g., Alzheimer’s disease). It is widely appreciated, as an example, that exogenously administered IL-2 has neuromodulatory actions starting from neurotrophic effects on septohippocampal neurons in culture and neurotransmitter release from cholinergic neurons, to hippocampal long-term potentiation and age-related changes in learning and memory. The vast majority of our knowledge of IL-2’s actions on brain function and behavior, however, is based on studies that use exogenously administered IL-2 to make inferences about the function of the endogenous cytokine.
Materials and Methods:
Animals and congenic breeding:
All mice in this study were cared for in compliance with the NIH Guide for the Care and Use of Laboratory Animals. Mice were housed in microisolater cages under specific pathogen free conditions. For GFP expression studies of the IL-2 transgene, female B6.Cg-Tg (IL2-EGFP) 17Evr (IL2p8-GFP) mice were obtained from the Mutant Mouse Regional Resource Center (University of Missouri) and bred in colony with C57BL/6 mice originally obtained from Jackson Laboratories. Transgene positive offspring were identified by PCR analysis of tail DNA. PCR primers in the IL-2 proximal promoter (IL2-1F: 5′-CATCCTTAGATGCAACCCTTCC-3′) and the GFP coding sequence (GFP-1R: 5′-GCTGAACTTGTGGCCGTTTAC-3′) were used, amplifying a 830-bp product in transgene-positive mice. PCR conditions were as follows: 95°C, 5 min, then 32 cycles of 95°C, 30 s; 58°C, 30 s; 72°C, 45 s, followed by a final 5 min at 72°C, using an i cycler (BioRad).
Tissue preparation: Mice were anesthetized by intraperitoneal injection of a 0.5 mg/mL ketamine cocktail in a 3:3:1 ratio (ketamine/xylazine/acepromazine) and were perfused with 4% paraformaldehyde (PF). Brains were dissected, post-fixed in 4% PF for 2 hrs at room temperature, and cryoprotected in 30% sucrose overnight at 4°C. Tissue was snap frozen in isopentane and stored at −80°C.
Acoustic startle reactivity and prepulse inhibition (PPI):
Two SR-LAB test chambers (San Diego Instruments) were used to measure acoustic startle response and prepulse inhibition as described previously. Mice were placed in a small cylindrical enclosure (3.8×9.5 cm) located in a dark, ventilated chamber. A speaker located 30 cm above the cylinder delivered the background noise (65 dB), startle stimuli, and prepulse stimuli, all of which consisted of broadband white noise.
Statistical analysis: Analysis of variance (ANOVA), and repeated measures ANOVA were used to make comparisions between subject groups. Post-hoc comparisons of interest were performed using the Fisher’s least significant difference test.
Results: We first tested the specificity of the GFP antibody by preincubating the primary antibody with recombinant GFP prior to use in our staining protocol to quench its ability to bind the antigen in tissue. As seen in Figure 1, as expected, immune cells (e.g., T cells, dendritic cells) in the white pulp area of the spleen expressed GFP, and the specificity of the primary antibody was clearly demonstrated by pre-incubation with recombinant GFP. As can be seen in Figure 2, a representative photograph of the septum and red nucleus, in most cells expressing GFP in the brain, the reporter was co-localized to the pan-neuronal cell marker, NeuN. Figure 3 shows GFP-positive cells identified in the medial and lateral septum, the fastigial nucleus, and the interposed nucleus of the cerebellum. In all areas examined, there were only a few cells that stained positive for GFP but not NeuN. Those brain cells were morphologically and geographically identical to brain cells expressing both markers. Across brain regions, we were ready to determine relative expression of IL-2 in IL2p8-GFP transgenic mice. GFP expression was found throughout the rostral-caudal extent of the brain and brainstem of IL2p8-GFP transgenic mice in discrete nuclei, and with a wide range of staining intensity.
Note: This work is partly presented at 5th Global Experts Meeting on Parkinsons, Huntingtons and Movement Disorders Oct 30-31, 2019 Tokyo, Japan
PDFShare this article
Mental Disorders and Treatment received 556 citations as per Google Scholar report